적요
The composition of high-molecular-weight-glutenin subunits (HMW-GS) is a key determinant of wheat baking properties. These subunits are encoded by the GLU-A1, GLU-B1, and GLU-D1 loci on the long arm of chromosome 1 and consist of x- and y-type subunits. Allelic variations in composition are a major factor influencing bakery quality. Unlike sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) or ultra-performance liquid chromatography (UPLC), which often fail to resolve closely related allelic variants, PCR-based markers allow for clear and definitive discrimination at the DNA level. Building on the results of a previous study that determined the GLU-B1 allele composition, we aimed to confirm—through the use of PCR markers—the allele compositions of GLU-A1 and GLU-D1 in 44 domestic wheat varieties. The results showed that “Jonong” and “Sinmichal1” contained the Glu-A1b (A1x2*) allele rather than Glu-A1a (A1x1) or Glu-A1c (A1x-null). Additionally, “Jonong” and “Sinmichal1” exhibited the allelic composition Glu-D1a (D1x2+D1y12), rather than Glu-D1d (D1x5+D1y10) or Glu-D1f (D1x2.2+D1y12). These results were compared with those obtained by SDS-PAGE and UPLC. The PCR-based markers used to identify GLU-A1 and GLU-D1 alleles in this study will be valuable for determining the allelic composition at the GLU-A1 and GLU-D1 loci in domestic wheat varieties. Furthermore, the re-evaluated genetic composition is expected to improve the precision of assessments related to the baking quality of domestic wheat.
서언
밀(
Triticum aestivum L.)은 다양한 식품의 밀가루 공급원으로, 밀가루가 다양한 식품으로 가공되는 능력은 주로 글루텐 단백질에 의해 결정된다. 성숙한 밀 종자는 일반적으로 8-20%의 단백질을 포함하고 있으며, 밀의 단백질 함량과 그 구성은 품질을 결정하는 중요 매개변수로 사용된다. 글루테닌과 글리아딘은 밀 글루텐의 주요 구성성분으로, 각각 글루텐 단백질의 약 40%와 60%를 차지한다. 글루테닌과 글리아딘 조성에 따라 결정되는 글루텐 강도와 신장성은 품질을 결정하는 주요 요소이다(
Brankovic et al. 2018,
Heo & Sherman 2013,
Shewry et al. 1992). 글루테닌 단백질은 고분자량 글루테닌 서브유닛(high-molecular-weight glutenin subunits, HMW-GS)과 저분자량 글루테닌 서브유닛(low-molecular-weight glutenin subunits, LMW-GS)으로 구성되며(
Gianibelli et al. 2001,
Lee et al. 2018,
Shewry et al. 1992), HMW-GS는 염색체 1번 장완에 위치한
GLU-
A1,
GLU-
B1,
GLU-
D1 유전자좌에 의해 암호화된다. 이들 유전자좌는 각각 x-형과 y-형 대립유전자로 구성된다(
Skerritt 1998).
GLU-
A1,
GLU-
B1,
GLU-
D1 유전자좌에서는 모두 광범위한 다형성이 발견되었으며,
GLU-B1과
GLU-D1 유전자좌는
GLU-A1 유전자좌보다 높은 다형성을 보인다. HMW-GS의 광범위한 대립유전자 다양성은 2020년 밀 유전자 카탈로그(WGC)에
GLU-
A1,
GLU-
B1 및
GLU-
D1에 대해 각각 53, 83 및 74개의 대립유전자가 보고되었다가 2024년 현재 각각 54, 88 및 75개로 업데이트되었다(
Andrade et al. 2024,
McIntosh 2024,
McIntosh et al. 2020). 일반밀 에서는 13개의
GLU-
A1 대립유전자(
Glu-
A1a,
Glu-
A1b,
Glu-
A1c,
Glu-
A1d,
Glu-
A1e,
Glu-
A1g,
Glu-
A1k,
Glu-
A1l,
Glu-
A1t,
Glu-
A1u,
Glu-
A1y,
Glu-
A1ba,
Glu-
A1bg)가 확인되었다. 그 중에
Glu-
A1x형 유전자는 주로 네 가지 대립유전자, 즉
Glu-A1a (A1x1),
Glu-A1b (A1x2*),
Glu-A1c (A1x-null)과
Glu-A1v (A1x2.1*)를 보유한 품종이 많으며,
Glu-A1y형 유전자는 특히 6배체 빵밀에서 발현이 억제되는 경우가 많은데, 이러한 침묵 현상은 주로 유전자 내에 존재하는 조기 종결 코돈으로 인해 단백질 번역이 중단되기 때문이다(
Payne & Lawrence 1983,
Shewry et al. 1992,
Thompson et al. 1983).
GLU-
A1,
GLU-
B1 및
GLU-
D1의 x-형 및 y-형 서브유닛을 식별하기 위한 프라이머 쌍과 특정 대립유전자를 식별하기 위한 프라이머 쌍은 1980년대 후반과 1990년대 초반에 보고되기 시작하였다(
Anderson & Greene 1989,
Anderson et al. 1989, D’
ovidio & Anderson 1994, D’
ovidio et al. 1994, 1995,
Halford et al. 1987). 기존에 보고된
GLU-A1,
GLU-B1,
GLU-D1 유전자좌의 대립유전자 판별 마커들과 최근에 추가된 판별 마커들로 다양한 대립유전자를 PCR 기반 마커를 통해 식별할 수 있게 되었다(
Ahmad 2000,
Butow et al. 2003,
Lee et al. 2024a,
Lee et al. 2025,
Lei et al. 2006,
Liu et al. 2008,
Ma et al. 2003,
Ragupathy et al. 2008,
Schwarz et al. 2004,
Smith et al. 1994,
Xu et al. 2008).
최근 국내에서 육성하여 재배한 44개 품종의 HMW-GS
Glu-B1x와
Glu-B1y 대립유전자 조성을 평가한 결과에서는 19개의 품종에서
Glu-B1x7 대립유전자가
Glu-B1x7*로, 2개의 품종에서
Glu-B1y8 대립유전자가
Glu-B1x8*로 판별되었다(
Lee et al. 2024b). 그리고 국내 밀 세 품종(조농, 신미찰1, 수강)의
GLU-
A1과
GLU-
D1의 대립유전자가 sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), two-dimensional gel electrophoresis (2-DE), reversed-phased high-performance liquid chromatography (RP-HPLC)와 matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-MS), multiplex PCR 분석결과에서 다르게 보고되었다(
Choi et al. 2022,
Jang et al. 2017, 2020,
Lee et al. 2013,
Park et al. 2011,
Shin et al. 2012). SDS-PAGE, RP-HPLC, MALDI-TOF-MS 분석에서 조농과 신미찰1은
Glu-A1c (A1x-null)과
Glu-
D1a (D1x2+D1y12)로 판별되었고(
Jang et al. 2017,
2020), 2-DE와 SDS-PAGE 분석에서 조농은
Glu-A1a (A1x1)와
Glu-
D1d (D1x5+D1y10), 신미찰1은
Glu-A1c (A1x-null)과
Glu-
D1f (D1x2.2+D1y12)로 판별되었다(
Lee et al. 2013).
기존의 단백질 분석 방법은 밀의 HMW-GS 대립유전자를 판별하는 데 널리 사용되어 왔지만 여러 가지 내재적 한계를 지닌다. SDS-PAGE에서는 분자량이 유사한 단백질이 서로 겹치는 이동 거리를 나타내어, 크기가 근소하게 다른 대립유전자를 명확히 구분하기 어렵다. 마찬가지로, UPLC 분석에서도 단백질 이소형 간의 머무름 중첩으로 인해 피크 분리가 불명확해지고, 결과 해석이 복잡해질 수 있다. 더욱이 두 방법 모두 시각적 판독과 분석자의 경험에 의존하기 때문에 대립유전자 식별 과정에서 일정 수준의 주관성이 개입될 가능성이 있다. 이에 반해, PCR기반 마커 분석은 HMW-GS를 발현하는 특정 DNA 염기서열을 직접 표적으로 하여, 단백질 기반 방법으로는 구별이 어려운 대립유전자 변이를 정확하고 재현성 있게 식별할 수 있다.
따라서 PCR 기반 유전자형 분석은 기존 단백질 분석법을 보완하는 강력한 접근법으로서, 밀 품질 평가에서 대립유전자 식별의 정확성과 신뢰성을 향상시키는 데 기여할 수 있다. 이에 본 연구에서는 최근에 보고된 PCR 기반 마커를 활용하여, 국내에서 육성된 44개 밀 품종의 GLU-A1과 GLU-D1의 대립유전자 조성을 평가하고자 하였다.
재료 및 방법
공시 재료
본 연구에서 이용된 밀 품종은 국내에서 육성 및 재배한 44개 품종으로 국립식량과학원(전라북도 완주군) 전작 포장에서 농촌진흥청 표준재배법(RDA 2012)에 준하여 재배한 후 수확하였다. 44개 품종 정보는
Lee et al. (2024b)에 보고되었으며
Table 1에 나타내었다. 유전자좌와 대립유전자는
Boden et al. (2023)이 보고한 밀 유전자 명명 지침에 따라 명명되었다.
Genomic DNA 추출은
Lee et al. (2024b)이 보고한 방법에 따라 수행하였다. PCR 반응에는 100 ng의 Genome DNA, 각 10 pmole의 forward 프라이머 및 reverse 프라이머, 1 x PCR master mix solution (인터론바이오, Korea)을 사용하였다. 프라이머 정보는
Table 2에 나타내었다. PCR조건은 94℃에서 5분간 pre-denaturation을 수행한 후 94℃에서 30초간 denaturation, 60-68℃ 사이의 조건에서 30초간 annealing, 72℃에서 30초간 extension 과정을 30회 반복하였다. 이후 final extension을 72℃에서 5분간 진행하였다. Annealing 온도와 밴드크기는
Table 2에 표시된 바와 같다. PCR 산물은 1.2% agarose gel에서 전기영동 후, 젤 이미지 시스템(Davinch-K, Korea)을 이용하여 이미지를 획득하였다(
Lee et al. 2022,
2023). PCR 분석은 2번 이상 반복하였다.
밀 글루테닌 단백질 분석은
Zhang et al. (2008)의 방법을 변형하여 밀 종실 1립에서 수행하였다. UPLC 분석은 Waters Alliance e2695 UPLC system (Alliance e2695, Waters Corp., MA, USA)을 사용하였으며,
Lee et al. (2024b)에서 보고한 방법에 따라 진행하였다. 분석에는 ACQUITY UPLC Peptide BEH C18 컬럼(300A, 1.7 μm, 2.1 mm×50 mm)과 photodiode array detector를 사용하였다. 이동상 용매는 0.1% trifluoroacetic acid를 포함한 H
2O (용매 A)와 0.1% trifluoroacetic acid를 포함한 acetonitrile (용매 B)를 사용하였다. 샘플 주입량은 3 μL, 유속은 0.55 μL/min으로 설정하였다. 용매 기울기는 0분에서 30분 동안 21% (B)에서 47% (B)로 변화시켰으며, 컬럼 온도와 시료 온도는 각각 55℃와 10℃로 유지였다.
글루테닌 단백질은 종실 1립에서 추출한 후, Bradford assay 용액과 반응시켜 Multiskan Sky High Microplate spectrophotometer (Thermo Scientific, USA)를 사용하여 595 nm에서 흡광도를 측정하였다. 단백질 약 10 μg을 4% stacking 젤에서 stacking 한 후, 10% separation 젤에서 분리하였다. 전기영동 후 젤은 Coomassie Brilliant Blue R+250 (Bio-Rad, USA) 염색용액으로 염색하고, 탈색용액 Ⅰ(10% acetic acid, 50% acetic acid, 50% methanol, 40% 증류수)과 탈색용액 Ⅱ(7% acetic acid, 5% methanol, 88% 증류수) 에서 순서대로 탈색시켜 단백질 밴드 패턴을 금강 품종과 비교하여 분석하였다(
Lee et al. 2024b).
결과 및 고찰
국내 밀 품종의 Glu-A1x 대립유전자 조성 평가
국내 밀 44개 품종의
Glu-A1x의 대립유전자 조성을 PCR 마커를 이용해 평가하고자 하였다. 기존에 보고된 국내 밀 44개 품종의 HMW-GS
GLU-A1,
GLU-B1,
GLU-D1 대립유전자 조성과 최근 수정된 대립유전자 조성을
Table 1에 제시하였으며, 사용한 프라이머(Primer set, PS) 정보와 PCR 조건은
Lee et al. (2025)의 연구를 참고하여
Table 2에 나타내었다.
이전연구에서 SDS-PAGE와 UPLC 분석을 통해
GLU-A1 대립유전자좌에서 A1x1과 A1x2*이 구분되지 않았으며, PCR 마커 분석 결과 일부 해외 품종에서 판별오류를 발견하였다(
Lee et al. 2025). 이전에 보고된 연구 결과에 따르면
Glu-A1x 대립유전자 판별에서 조농, 신미찰1, 수강에 대해 세 연구 그룹에서 다르게 평가된 바 있다.
Jang et al. (2017,
2020)은 SDS-PAGE, RP-HPLC와 MALDI-TOF-MS 분석을 통해 조농과 신미찰1의
GLU-
A1 대립유전자 조성은
Glu-A1c (A1x-null), 수강은
Glu-A1b (A1x2*)로 보고되었다. 또한, 2-DE와 SDS-PAGE 결과에서 조농은
Glu-A1a (A1x1), 수강은
Glu-A1b (A1x2*)로 보고되었고, 신미찰1은
Glu-A1c (A1x-null) 또는
Glu-A1b (A1x2*)로 보고되었다(
Lee et al. 2013,
Park et al. 2011,
Shin et al. 2012).
Choi et al. (2022)은 multiplex-PCR 분석을 통해 세 품종 모두를
Glu-A1c (A1x-null)로 보고하였다. 따라서 본 연구에서는 국내 밀 44개 품종의
Glu-A1x의 대립유전자 조성을 PCR 기반 마커(PS1-PS4,
Table 2)로 분석하고자 하였다.
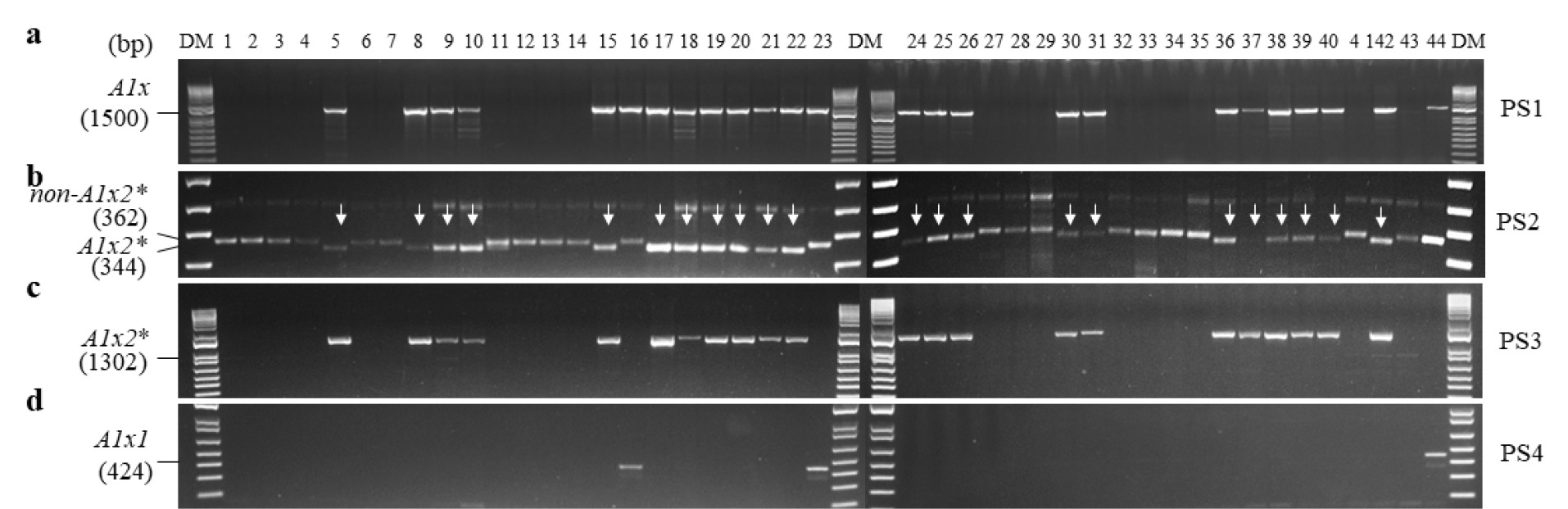
국내 밀 44개 품종 중에서 올, 그루, 다홍, 청계, 남해, 우리, 서둔, 새올, 진품, 밀성, 은파, 고분, 안백, 황금알, 태중, 조은, 조품, 조중, 중모2008은
Glu-A1x 대립유전자가
Glu-A1x-
null로 판별되었으며, 나머지 25개 품종은
Glu-A1x-
null이 아닌 것으로 판별되었다(PS1,
Fig. 1A).
Glu-A1x-
null이 아닌 25개 품종 중 탑동, 올그루, 알찬, 금강, 신미찰, 다분, 한백, 수안, 고소, 조아, 호중, 새금강, 아리흑, 백찰, 조농, 신미찰1, 연백, 백중, 적중, 수강, 다중, 조한은
Glu-A1x2*를 포함하고 있었다(PS2-3,
Figs. 1B,
1C). 나머지 세 품종 조경, 백강, 우주는 PS4에서
Glu-
A1x1 특이적으로 검출되었다(
Fig. 1D). 본 연구에서 PCR 분석 결과,
Glu-A1a (A1x1) 또는
Glu-A1c (A1x-null)로 보고된 조농과
Glu-A1b (A1x2*) 또는
Glu-A1c (A1x-null)로 보고된 신미찰1과 수강은 모두
Glu-A1b (A1x2*)를 보유한 것으로 판별되었다(
Fig. 1,
Table 1).
44개 품종 중에 조농과 신미찰1의
GLU-D1 대립유전자 조성은 여러 연구에서 다르게 보고되었다. RP-HPLC와 MALDI-TOF-MS 분석결과에서는
Glu-
D1a (D1x2+D1y12)로 판별되었고(
Jang et al. 2017,
2020), 2-DE와 SDS-PAGE분석에서는 조농이
Glu-
D1d (D1x5+D1y10), 신미찰1이
Glu-
D1f (D1x2.2+D1y12) 또는
Glu-
D1a (D1x2+D1y12)로 판별되었다(
Lee et al. 2013,
Park et al. 2011,
Shin et al. 2012). 본 연구에서는 먼저 국내 밀 44개 품종의
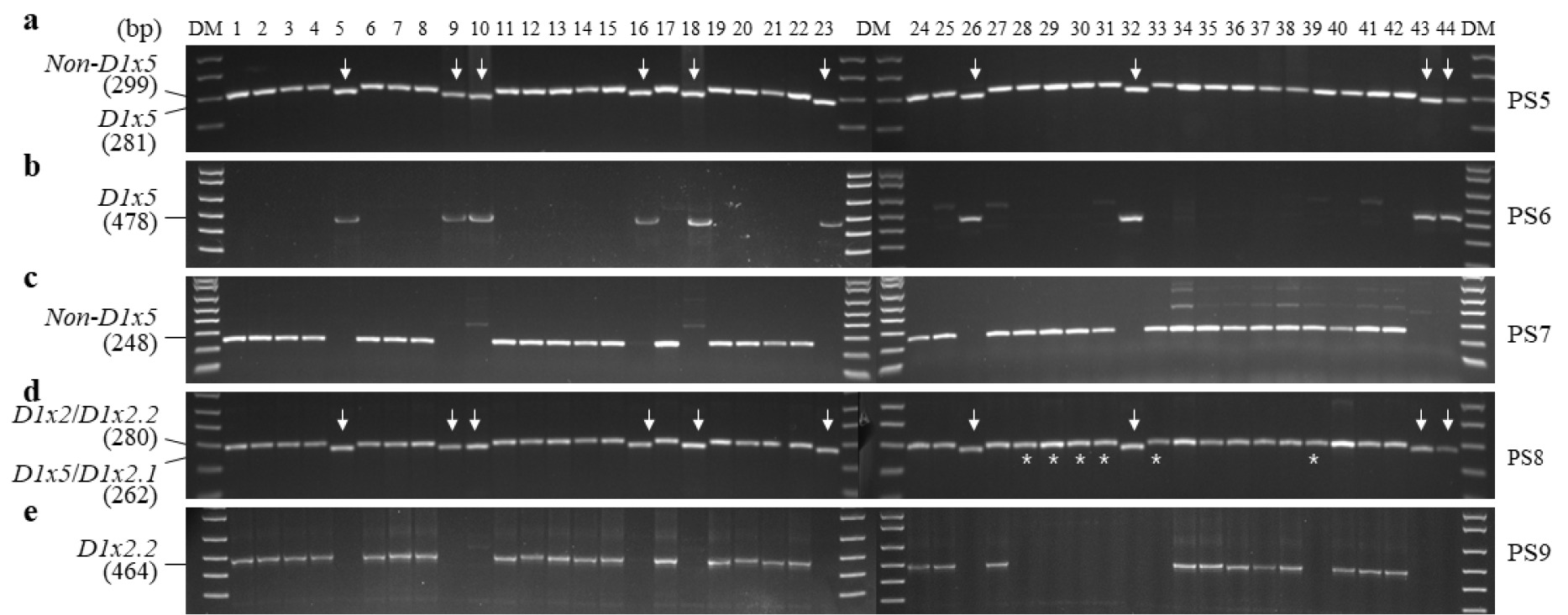
Glu-D1x 대립유전자 조성을 PCR 마커를 이용해 평가하였다(
Fig. 2). PS5을 사용하여
Glu-
D1x5와
non-
Glu-
D1x5를 구분한 결과, 탑동, 알찬, 금강, 조경, 한백, 백강, 백찰, 황금알, 중모2008, 우주가
Glu-
D1x5로 판별되었으며, PS6에서도
Glu-
D1x5 특이적으로 검출되었다(
Figs. 2A,
2B). PS7을 사용한 분석에서는 나머지 34개 품종이
non-
Glu-
D1x5 특이적으로 검출되는 것을 확인하였다(
Fig. 2C). PS8에서는 34개 품종이
Glu-
D1x2 또는
Glu-
D1x2.2로 판별되었으며, 나머지 10개 품종은
Glu-
D1x5 또는
Glu-
D1x2.1로 판별되었다.
Glu-
D1x5 또는
Glu-
D1x2.1로 판별된 품종들은 PS5와 PS6에서도 모두
Glu-
D1x5로 판별되었으므로,
Glu-
D1x2.1 대립유전자를 포함하는 품종은 없는 것으로 확인되었다(
Fig. 2D).
Glu-
D1x2 또는
Glu-
D1x2.2로 구분된 34개 품종 중에 올, 그루, 다홍, 청계, 남해, 우리, 올그루, 서둔, 새올, 진품, 밀성, 신미찰, 다분, 수안, 고소, 조아, 호중, 새금강, 아리흑, 은파, 조은, 조품, 연백, 백중, 적중, 다중, 조중, 조한은 PS9에서
Glu-
D1x2.2 특이적인 마커로 검출되었으며, 나머지 고분, 안백, 조농, 신미찰1, 태중, 수강은
Glu-
D1x2 대립유전자로 판별되었다(
Figs. 2D,
2E).
다음으로, 국내 밀 44개 품종의
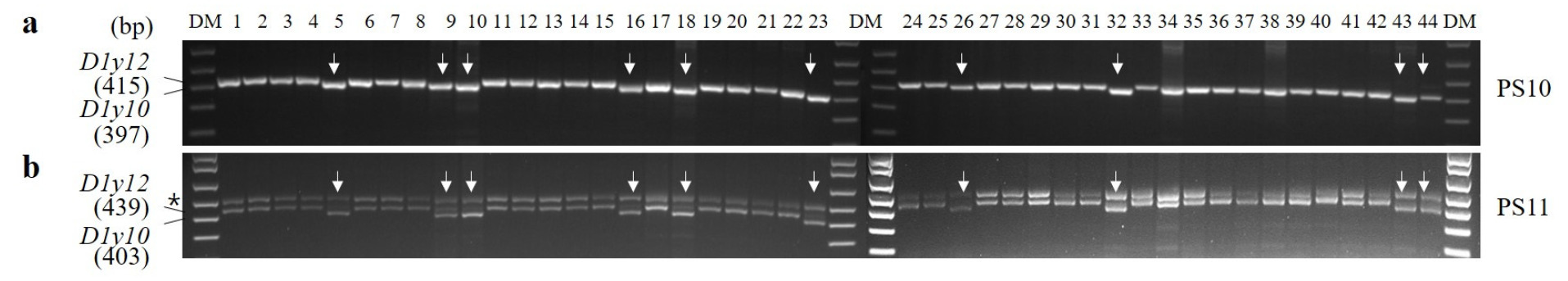
Glu-D1y 대립유전자 조성을 PCR 마커로 평가하였다. PS10과 PS11을 사용하여
Glu-D1y10과
Glu-D1y12를 구분한 결과, 탑동, 알찬, 금강, 조경, 한백, 백강, 백찰, 황금알, 중모2008, 우주가
Glu-D1y10으로 판별되었으며, 나머지 34개 품종은
Glu-D1y12로 판별되었다(
Fig. 3). 국내 밀 44개 품종 중에 10개 품종은
Glu-D1d (D1x5+D1y10), 6개 품종은
Glu-D1a (D1x2+D1y12), 28개 품종은
Glu-D1f (D1x2.2+D1y12)의 조성임을 알 수 있었다. 따라서 본 연구에서 PCR 마커로 분석한 결과에서 조농과 신미찰1의 대립유전자 조성은
Glu-
D1a (D1x2+D1y12)로 확인되었다(
Fig. 2).
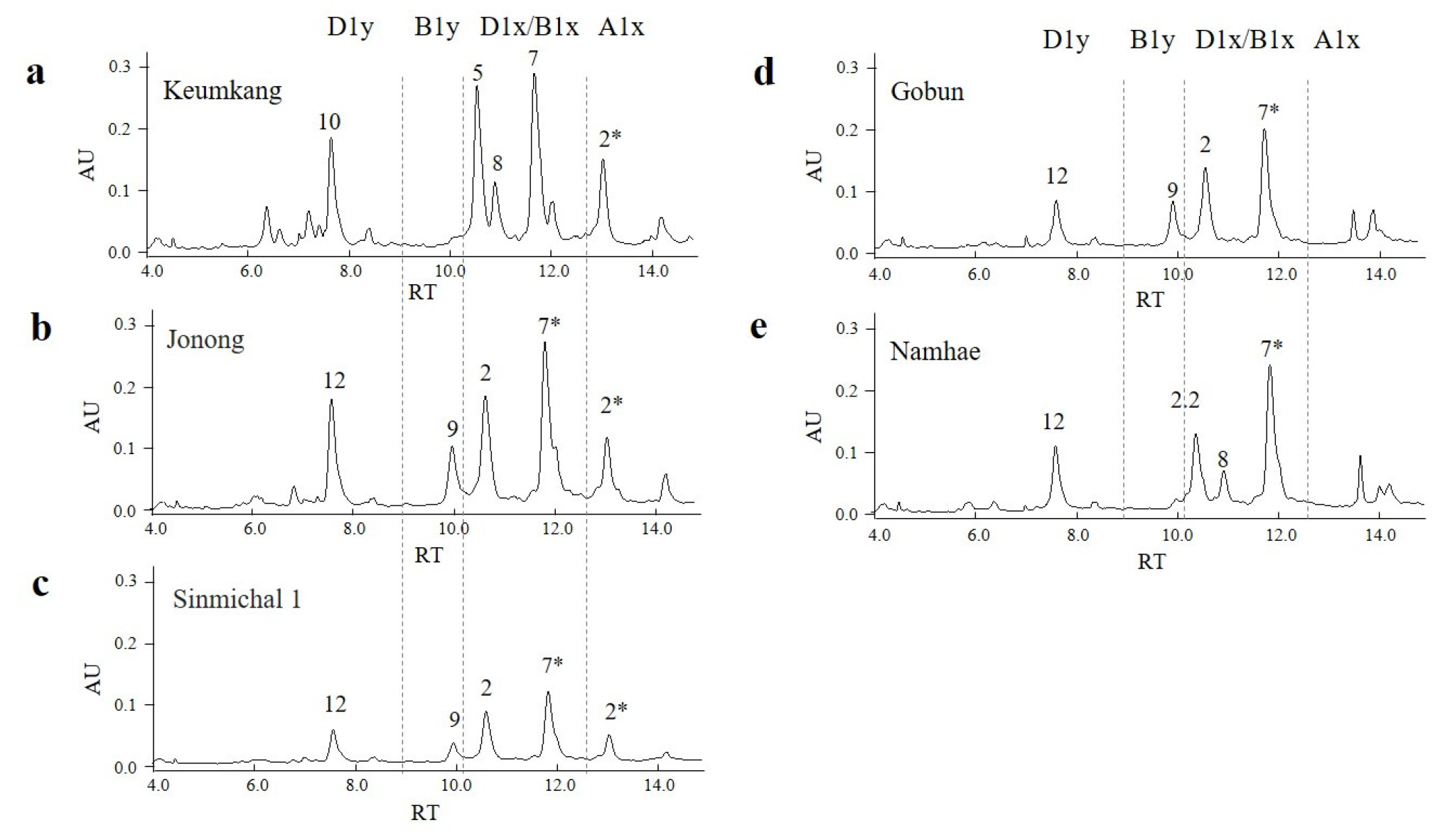
UPLC를 이용하여 조농과 신미찰1의 HMW-GS 조성을 분석하였으며(
Fig. 4), 금강, 고분, 남해 품종을 비교 품종으로 사용하였다. 조농과 신미찰1에서는 금강과 유사한 위치에서 A1x 피크가 검출되었다. 이전 연구에 따르면 A1x1 (Ranee)과 A1x2*(금강, 탑동, 신미찰1)의 평균 용출 시간은 각각 13.05±0.05분과 13.08±0.02분으로 오차범위가 각각 0.09분과 0.07분에 불과하여 용출 시간만으로 두 단백질을 구분하기는 어려웠다(
Lee et al. 2024a,
2024b)). 본 연구에서 조농과 신미찰1의 A1x의 용출시간은 각각 13.07±0.01분과 13.10±0.01 분으로, 금강의 13.06±0.01과 큰 차이를 보이지 않았으며 A1x1과 A1x2*의 오차 범위 네에 포함되었다.
D1x2.2 (올, 청계, 은파, 남해, 서둔, 진품), D1x5 (금강, 탑동, 백찰), D1x2 (고분, 안백, 태중)의 용출 시간은 각각 10.39±0.01, 10.55±0.01, 10.65±0.02분으로 D1x2.2, D1x5, D1x2 순으로 분리되었다(
Lee et al. 2024b). 조농과 신미찰1의 D1x의 용출시간은 각각 10.63±0.01과 10.66±0.02분으로 D1x5 용출시간보다 다소 늦게 나타났으며, 이전 연구에서 보고된 D1x2의 추출시간과 유사한 수준이었다. 한편, D1y10 (금강, 탑동)과 D1y12 (올, 청계)의 추출시간은 7.64±0.01과 7.61±0.01로 거의 차이가 없었다(
Lee et al. 2024b). 본 연구에서 조농과 신미찰1의 D1y의 추출시간은 각각 7.57±0.01과 7.62±0.01분으로, D1y10의 용출시간(7.64±0.01분)과 유사하였다(
Fig. 4). 이러한 결과는 UPLC 분석만으로는 A1x1과 A1x2*, D1y10과 D1y12의 구분이 명확하지 않아, 대립유전자 판별의 정확성에 한계가 있음을 시사한다.
다음으로, SDS-PAGE 분석을 통해 조농과 신미찰1의 단백질 조성을 금강 품종과 비교하였다(
Fig. 5). A1x2*와 A1x1은 각각 815와 830 아미노산으로 구성되며, 이들 예상 분자량은 각각 88.56 kDa과 89.90 kDa이다. 실제 SDS-PAG 결과에서 100 kDa 부근에서 밴드가 검출되었다(
Lee et al. 2025). 본 연구에서도 금강, 조농, 신미찰1 모두에서 100 kDa 근처에서 유사한 크기의 밴드가 확인되었다. 이전 연구에 따르면, D1x2.2, D1x2, D1x5는 각각 971, 843, 838개의 아미노산으로 구성되며, 예상 분자량은 각각 103.15, 90.38, 89.26 kDa이다. 이 중 D1x2.2는 상대적으로 분자량이 커서 SDS-PAGE에서 이동 속도가 느렸고, 실제로 135 kDa 마커 아래에서 검출되어 다른 단백질들과 명확히 구별되었다. 그러나 D1x2와 D1x5는 100 kDa 부근에서 검출되었으며, 크기 차이가 작고 A1x2*와 이동 속도가 유사하여 상호 구별이 어려웠다. 한편, D1y10과 D1y12는 각각 648개와 660개의 아미노산으로 구성되며, 이들의 분자량은 각각 69.69 kDa과 70.93 kDa이다. SDS-PAGE상에서는 두 단백질 모두 75 kDa 마커 근처에서 검출되었다(D’
ovidio et al. 1994,
Lee et al. 2025). 조농과 신미찰1의 D1y12는 금강의 D1y10보다 약간 느리게 이동하는 경향을 보였다. 이러한 결과는 SDS-PAGE상에서 단백질의 이동 속도가 유사한 경우, 단백질 조성을 정확하게 판별하기 어려울 수 있음을 시사한다.
Jang et al. (2020)이 수행한 MALDI-TOF-MS 분석에서는 A1x2*가 D1x2와 부분적으로 중첩되어 관찰되었으며, 이에 따라 해당 대립유전자가 분석 결과에서 제외된 것으로 판단된다. 그리고,
Jang et al. (2017)의 SDS-PAGE 결과에서도 A1x2*와 D1x2 단백질의 밴드가 부분적으로 중첩되어 나타났을 가능성이 있으며, 이로 인해 명확한 구분이 어려웠을 것으로 추정된다. 더불어
Lee et al. (2013)이 수행한 2-DE 분석에서도 유사한 현상이 나타났을 가능성이 있다. 이와 같이 SDS-PAGE 및 2-DE는 단백질 간 분자량이나 등전점이 유사한 경우 밴드 또는 스팟이 서로 겹쳐 나타나기 쉬우며, 이는 동일한 또는 유사한 이동거리⋅위치로 인해 대립유전자 구분의 정확성을 저하시킬 수 있다. 마찬가지로, UPLC와 같은 액체크로마토그래피 분석에서도 단백질 이소형 간 머무름(retention time)의 중첩이 발생할 수 있어, 피크 분리가 불명확해지고 결과 해석에 주관성이 개입될 가능성이 있다. 이러한 단백질 기반 분석법의 한계는 밀의 HMW-GS 대립유전자 구성을 판별하는 과정에서 오류나 불일치가 발생할 가능성을 높인다.
본 연구에서는 기존 SDS-PAGE 또는 UPLC 분석에서 구분이 불명확했던 A1x1, A1x2*, D1x2, D1y12 등의 대립유전자가 PCR 분석을 통해 명확하게 구분되었으며, 이는 단백질 기반 분석법의 한계를 효과적으로 보완할 수 있음을 보여준다. 따라서 PCR 기반 마커 분석은 국내 밀 품종의 HMW-GS 대립유전자 구성을 명확히 규명하고, 제빵 품질 관련 유전적 특성을 보다 정밀하게 평가하는 데 유용한 도구로 활용될 수 있을 것으로 판단된다.
국내 밀 품종의 HMW-GS 대립유전자 조성
새로 판별한 국내 밀 44개 품종의 HMW-GS
GLU-
A1과
GLU-
D1의 대립유전자 유전자 조성을
Table 1에 나타내었다. 국내 밀의
Glu-
A1x 조성은
Glu-
A1a (A1x1)가 3개 품종,
Glu-
A1b (A1x2*)는 22개 품종에서 확인되었으며
Glu-
A1c (A1x-null)는 19개 품종에서 확인되었다. 국내 밀의
GLU-
A1 대립유전자 조성은 주로
Glu-
A1b (A1x2*)와
Glu-
A1c (A1x-null)로 구성되어 있었다. 그리고 국내 밀 44개 품종의
GLU-
D1 대립유전자 조성은 세 가지 대립유전자 조성으로 이루어졌으며,
Glu-
D1d (D1x5+D1y10)이 10개 품종,
Glu-
D1a (D1x2+D1y12)는 6개 품종,
Glu-
D1f (D1x2.2+D1y12)는 28개 품종에서 확인되었다.
결론
국내에서 육성된 44개 밀 품종 중에 조농, 신미찰1, 수강의 HMW-GS GLU-A1과 GLU-D1 대립유전자는 기존 단백질 분석법에서 일부 판별 오류가 확인되었다. PCR 마커 분석 결과, 세 품종의 Glu-A1x는 모두 Glu-A1b (A1x2*)로, 조농과 신미찰1의 GLU-D1은 Glu-D1a (D1x2+D1y12)로 정확히 판별되었다. 본 연구는PCR 기반 분석을 통해 국내 밀 HMW-GS 유전자좌의 대립유전자를 간편하고 정확하게 판별할 수 있음을 보여주었으며, 이는 국내 밀 품종등록과 제빵품질 예측 평가의 표준화에도 기여할 수 있는 실용적 도구로 활용될 수 있다.
적요
HMW-GS의 조성은 단백질 특성과 제빵특성에 중요한 역할을 하며, HMW-GS는 1번 염색체 장완에 위치한 GLU-A1, GLU-B1, GLU-D1 유전자좌에 의해 암호화되며, x-형과 y-형의 대립유전자로 구성된다. HMW-GS 구성의 대립형질 변이는 제빵 품질에 영향을 주는 주요 요인이다. SDS-PAGE나 UPLC는 밀접하게 관련된 대립유전자의 변이를 구분하기 어려운 경우가 많지만, PCR 기반 마커는 DNA 수준에서 명확하고 신뢰성 있는 식별이 가능하다. 이전 연구에서 보고된 GLU-B1 대립유전자 조성의 PCR 마커를 이용한 판별에 이어, 본 연구에서는 국내 밀 44개 품종의 GLU-A1과 GLU-D1의 대립유전자 조성을 PCR 마커로 확인하고자 하였다. 44개 품종 중 조농, 신미찰1, 수강의 Glu-A1x 유전자 조성과 조농과 신미찰1의 Glu-D1x 및 Glu-D1y 유전자 조성이 재평가되었다. 또한, PCR 마커 판별 결과 기존 보고와 다르게 판별된 품종들의 HMW-GS 단백질을 SDS-PAGE로 분석하여 비교하였다. 본 연구에서 사용된 PCR 기반 마커 정보는 국내 밀 품종의 GLU-A1과 GLU-D1의 대립유전자 조성 판별에 유용하게 사용될 것이며, 재평가된 유전자 조성은 국내 밀의 제빵 특성 평가에 정확성을 제공할 것으로 기대된다.
사사
본 연구는 2025년도 농촌진흥청 국립식량과학원 전문연구원 과정 지원사업과 농촌진흥청 연구사업(가공 용도별 밀 품종개발 및 육종효율 증진연구(4단계)(과제번호: PJ01677101)에 의해 이루어진 결과로 이에 감사드립니다.
Fig. 1PCR analysis of Glu-A1x alleles in 44 domestic wheat cultivars. DM, DNA size marker; PS, primer set. Arrows represent indels.
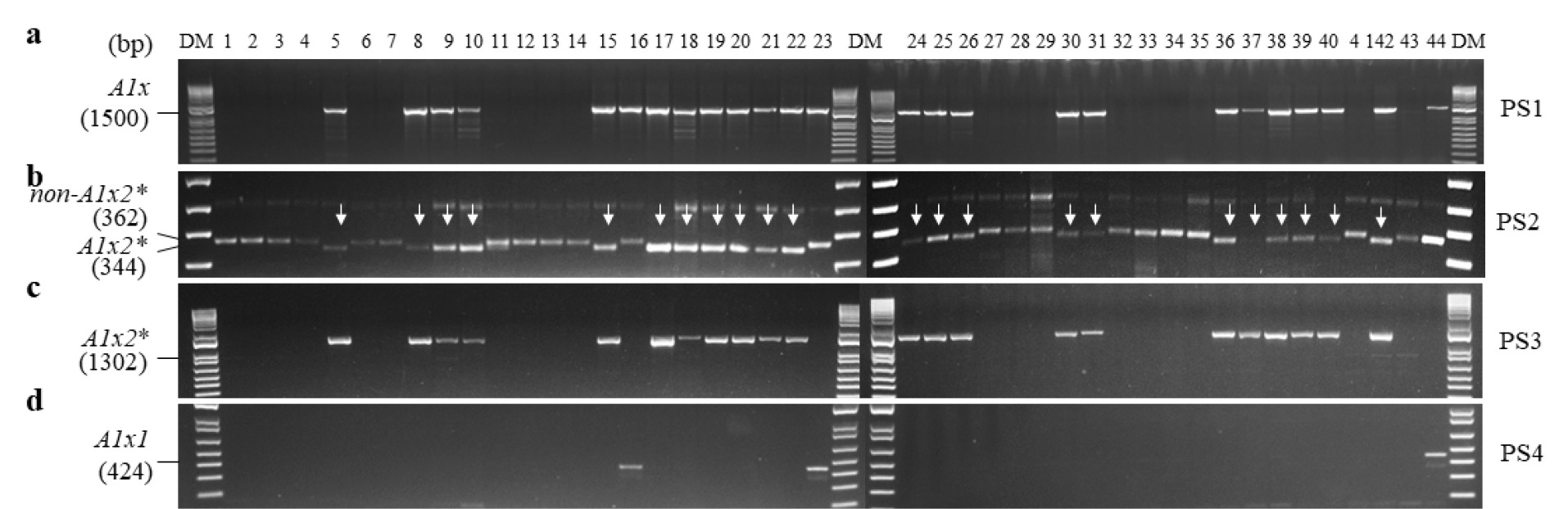
Fig. 2PCR analysis of Glu-D1x alleles in 44 domestic wheat cultivars. DM, DNA size marker; PS, primer set. Arrows represent indels, and the asterisk (*) denotes Glu-D1x2.
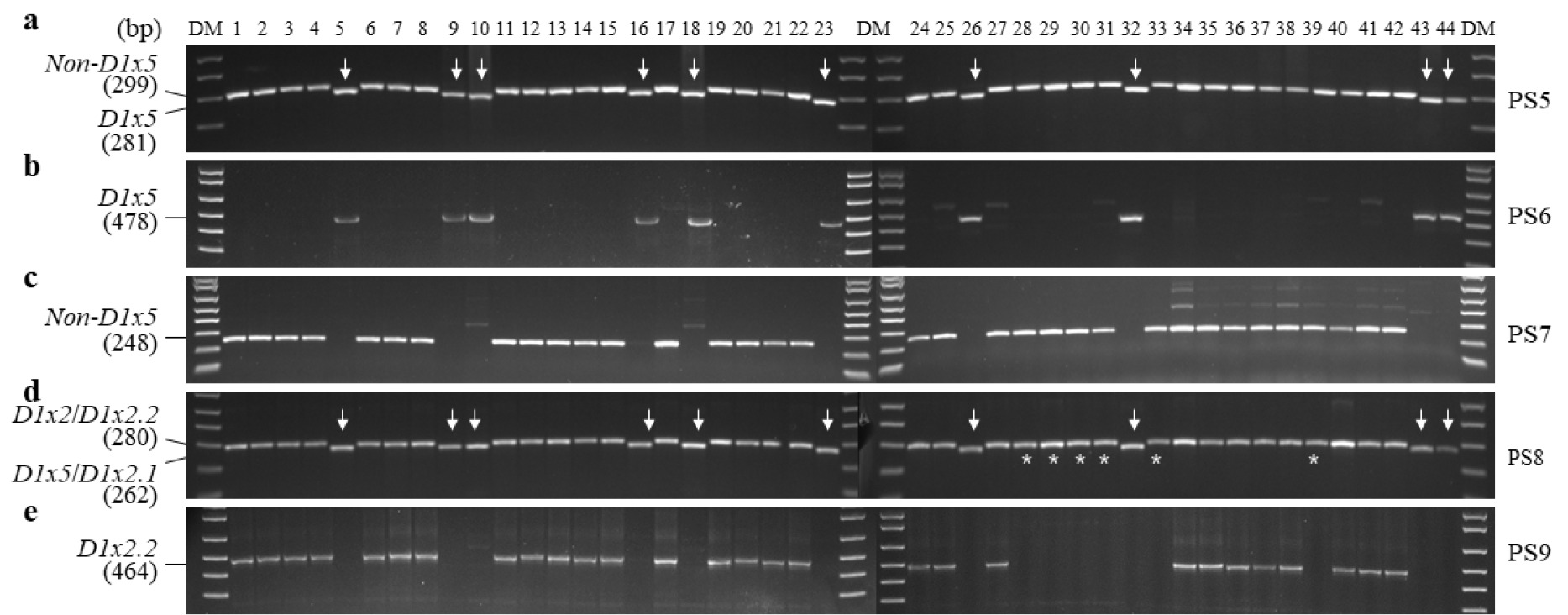
Fig. 3PCR analysis of Glu-D1y alleles in 44 domestic wheat cultivars. DM, DNA size marker; PS, primer set. Arrows represent indels.
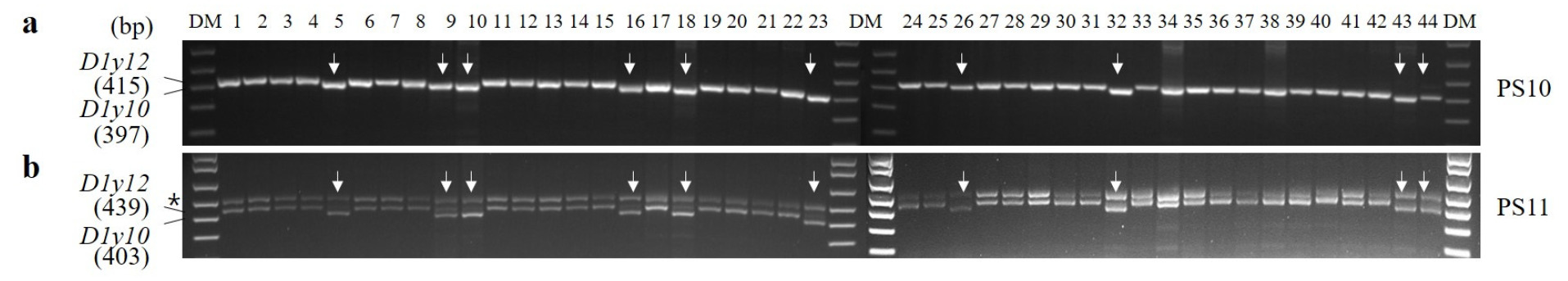
Fig. 4Expression of HMW-GSs in five domestic wheat cultivars were compared using Ultra Performance Liquid Chromatography. Retention times (RTs) were determined for each peak in the HMW-GS region. AU, arbitrary units. The numbers correspond to HMW-GS subunits as follows: 2* represent A1x2*; 7 and 7* represent B1x7 and B1x7*; 8 and 9 represent B1y8 and Bly9; 2,2, 2, and 5 represent D1x2.2, D1x2, and D1x5; 10 and 12 represent D1y10 and D1y12, respectively.
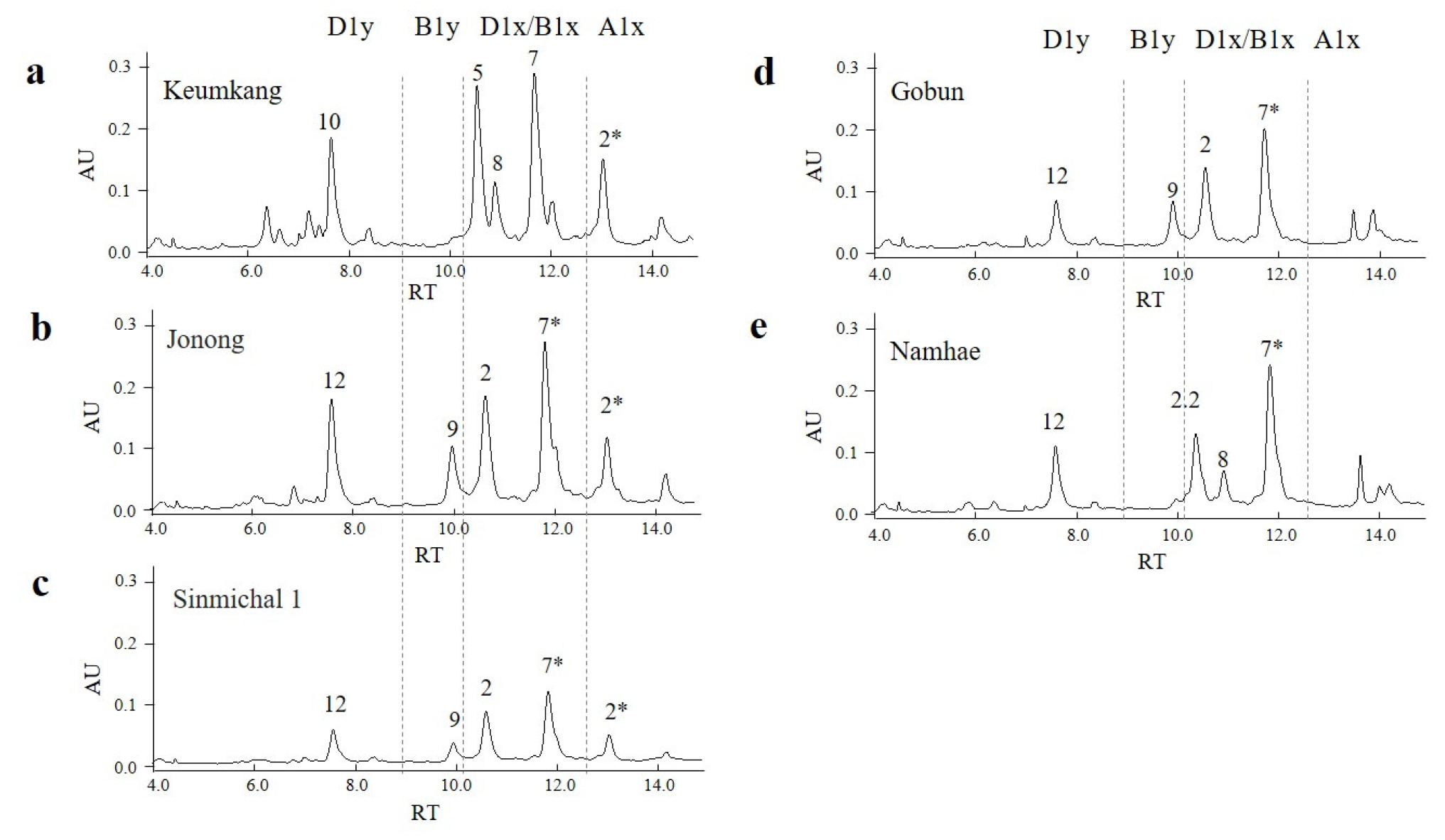
Fig. 5Protein composition of the HMW-GSs in two domestic wheat cultivars (Jonong and Sinmichal1) compared with Keumkang. PM, protein marker; HMW-GS, high-molecular-weight glutenin; LMW-GS, low-molecular-weight glutenin. The numbers correspond to HMW-GS subunits as follows: 2* represent A1x2*; 7 and 7* represent B1x7 and B1x7*; 8 and 9 represent B1y8 and Bly9; 2 and 5 represent D1x2 and D1x5; 10 and 12 represent D1y10 and D1y12, respectively.

Table 1The information of HMW-GS GLU-A1, GLU-B1, and GLU-D1 alleles in 44 domestic wheat cultivars.
Table 1
|
No. |
Name |
Pedigree |
Reported |
|
Corrected |
|
|
|
GLU-A1
|
GLU-B1
|
GLU-D1
|
GLU-A1
|
GLU-D1
|
|
|
|
|
|
|
x |
x+y |
x+y |
x |
x+y |
|
1 |
Ol |
Norin72/Norin12 |
N |
7*+8 |
2.2+12 |
|
N |
2.2+12 |
|
2 |
Geuru |
Strampelli/69D-3607//Chokwang |
N |
7+8 |
2.2+12 |
|
N |
2.2+12 |
|
3 |
Dahong |
Norin72/Wonkwang |
N |
7+8 |
2.2+12 |
|
N |
2.2+12 |
|
4 |
Chungkye |
Norin4/Sharbatisonora |
N |
7*+8 |
2.2+12 |
|
N |
2.2+12 |
|
5 |
Tapdong |
Chugoku81//Suwon158/Toropi |
2* |
7*+8 |
5+10 |
|
2* |
5+10 |
|
6 |
Namhae |
Ol/Calidad |
N |
7*+8 |
2.2+12 |
|
N |
2.2+12 |
|
7 |
Uri |
Geuru/Ol |
N |
7+8 |
2.2+12 |
|
N |
2.2+12 |
|
8 |
Olgeuru |
Geuru/Chokwang//Saikai143 |
2* |
7+8 |
2.2+12 |
|
2* |
2.2+12 |
|
9 |
Alchan |
Suwon210/Tapdong |
2* |
7+8 |
5+10 |
|
2* |
5+10 |
|
10 |
Keumkang |
Geuru/Kanto75//Eunpa |
2* |
7+8 |
5+10 |
|
2* |
5+10 |
|
11 |
Seodun |
Geuru/Genaro81 |
N |
7*+8 |
2.2+12 |
|
N |
2.2+12 |
|
12 |
Saeol |
Shirogane//Norin43/Sonalika |
N |
7+8 |
2.2+12 |
|
N |
2.2+12 |
|
13 |
Jinpoom |
Geuru/Genaro81 |
N |
7*+8 |
2.2+12 |
|
N |
2.2+12 |
|
14 |
Milsung |
Shirogane//Norin43/Sonalika |
N |
7+8 |
2.2+12 |
|
N |
2.2+12 |
|
15 |
Sinmichal |
Olgeuru//Kanto107/BaiHuo |
2* |
7*+8* |
2.2+12 |
|
2* |
2.2+12 |
|
16 |
Jokyung |
Seri82/Keumkang |
1 |
7+8 |
5+10 |
|
1 |
5+10 |
|
17 |
Dabun |
Suwon234//sw76039/Suwon22 |
2* |
7+8 |
2.2+12 |
|
2* |
2.2+12 |
|
18 |
Hanbaek |
Shann7859/Keumkang//Guamuehill |
2* |
7+8 |
5+10 |
|
2* |
5+10 |
|
19 |
Suan |
Keumkang/Eunpa//Keumkang |
2* |
7*+8 |
2.2+12 |
|
2* |
2.2+12 |
|
20 |
Goso |
Gobun/Ol |
2* |
7+8 |
2.2+12 |
|
2* |
2.2+12 |
|
21 |
Joa |
SW86054-MB-27-3-2-1-1-1/Sumai#3 |
2* |
7*+8 |
2.2+12 |
|
2* |
2.2+12 |
|
22 |
Hojoong |
Alchan2/3/Chunm18//JUP/BJY/4/Keumkang |
2* |
7*+8 |
2.2+12 |
|
2* |
2.2+12 |
|
23 |
Baekkang |
Topdong/Klasic |
1 |
7+8 |
5+10 |
|
1 |
5+10 |
|
24 |
Saekeumkang |
Keumkang/Olgeuru |
2* |
7+8 |
2.2+12 |
|
2* |
2.2+12 |
|
25 |
Ariheuk |
K253305/Sinmichal |
2* |
7*+8* |
2.2+12 |
|
2* |
2.2+12 |
|
26 |
Baekchal |
Sinmichal/Keumkang |
2* |
7*+8* |
5+10 |
|
2* |
5+10 |
|
27 |
Eunpa |
Chugoku81/3/Tob-CNO/Yuksung3/Suwon185 |
N |
7*+9 |
2.2+12 |
|
N |
2.2+12 |
|
28 |
Gobun |
Eunpa/Tapdong//Eunpa/Shannung6521 |
N |
7*+9 |
2+12 |
|
N |
2+12 |
|
29 |
Anbaek |
Sae/Geuru |
N |
7*+9 |
2+12 |
|
N |
2+12 |
|
30 |
Jonong |
Suwon234/SW80199 |
1zyx/Nwvu
|
7*+9 |
(5+10)zyxw/(2+12)v
|
|
2* |
2+12 |
|
31 |
Sinmichal1 |
Alchan//Kanto107/BaiHuo |
2*x/Nywvu
|
7*+9 |
(2+12)xwv/(2.2+12)y
|
|
2* |
2+12 |
|
32 |
Hwanggeumal |
Jokyung/Kauz/Rayo//Jopoom |
N |
7*+9 |
5+10 |
|
N |
5+10 |
|
33 |
Taejoong |
Xian83(104).11/Keumkang |
N |
7*+9 |
2+12 |
|
N |
2+12 |
|
34 |
Joeun |
Eunpa/Suwon242 |
N |
13+16 |
2.2+12 |
|
N |
2.2+12 |
|
35 |
Jopoom |
Kanto75//OR8500494P/Bezostaya |
N |
13+16 |
2.2+12 |
|
N |
2.2+12 |
|
36 |
Younbaek |
Keumkang/Tapdong |
2* |
13+16 |
2.2+12 |
|
2* |
2.2+12 |
|
37 |
Baekjoong |
Keumkang/Olgeuru |
2* |
13+16 |
2.2+12 |
|
2* |
2.2+12 |
|
38 |
Jeokjoong |
Keumkang/Tapdong |
2* |
13+16 |
2.2+12 |
|
2* |
2.2+12 |
|
39 |
Sukang |
Suwon266/Asakaze |
2*zywv/Nu
|
13+16 |
2+12 |
|
2* |
2+12 |
|
40 |
Dajoong |
SW992114-NM-131-7/Gobun |
2* |
13+16 |
2.2+12 |
|
2* |
2.2+12 |
|
41 |
Jojoong |
Suwon272/Olgeuru//Keumkang/Suwon252 |
N |
13+16 |
2.2+12 |
|
N |
2.2+12 |
|
42 |
Johan |
Suwon262/Joeun |
2* |
13+16 |
2.2+12 |
|
2* |
2.2+12 |
|
43 |
Joongmo2008 |
Eunpa*2//SH3/CBRD/3/Keumkang |
N |
17+18 |
5+10 |
|
N |
5+10 |
|
44 |
Wooju |
Jokyung/K253306 |
1 |
20+20 |
5+10 |
|
1 |
5+10 |
Table 2Primer information used for detection of GLU-A1 and GLU-D1 in present study. PS, primer set; A1x, Glu-A1x; D1x, Glu-D1x; D1y, Glu-D1y.
Table 2
|
PS |
Gene |
Primer name |
5'→3' |
AT (℃) |
Size (bp) |
Reference |
|
1 |
A1x/non-A1x
|
c |
CCATCGAAATGGCTAAGCGG |
68 |
1500 |
Lafiandra et al. (1997)
|
|
d |
GTCCAGAAGTTGGGAAGTGC |
|
2 |
A1x2*/non-A1x2*
|
UMN19-F |
CGAGACAATATGAGCAGCAAG |
66 |
344/362 |
Liu et al. (2008)
|
|
UMN19-R |
CTGCCATGGAGAAGTTGGA |
|
3 |
A1x2* |
Ax2*-F |
ATGACTAAGCGGTTGGTTCTT |
68 |
1302 |
Ma et al. (2003)
|
|
Ax2*-R |
ACCTTGCTCCCCTTGTCTTT |
|
4 |
A1x1
|
MHAX1-F |
TCCAACTTCTCCATGGCAG |
68 |
443 |
Lee et al. (2025)
|
|
MHAX1-R3 |
GCTGCTGCGCAGAAGTTGGA |
|
5 |
D1x5/non-D1x5
|
UMN25-F |
GGGACAATACGAGCAGCAAA |
68 |
281/299 |
Liu et al. (2008)
|
|
UMN25-R |
CTTGTTCCGGTTGTTGCCA |
|
6 |
D1x5
|
Dx-5-1-F |
CGTCCCTATAAAAGCCTAGC |
60 |
478 |
Liu et al. (2008)
|
|
Dx-5-1-R |
AGTATGAAACCTGCTGCGGAC |
|
7 |
Non-D1x5
|
MHDx-F |
GTCGCGGGACAATACGAGCG |
68 |
248 |
Lee et al. (2025)
|
|
MHDx-R2 |
CCTTGTTGTCCTTGTCCTGAT |
|
8 |
D1x5, D1x2.1/D1x
|
MHDX-F2 |
CCAACTTCTCCGCAGGAGTC |
68 |
262/280 |
Lee et al. (2025)
|
|
D1x2, D1x2.2
|
MHDX-R4 |
TTGCAGCCATTGTCCTGGCC |
|
9 |
D1x2.2
|
MHDX2.2-F3 |
TACTACCCAACTTCTCCGCT |
68 |
464 |
Lee et al. (2025)
|
|
MHDX2.2-R4 |
CTGCTGCGGAGAAGTTAGGT |
|
10 |
D1y10/D1y12
|
UMN26-F |
CGCAAGACAATATGAGCAAACT |
60 |
397/415 |
Liu et al. (2008)
|
|
UMN26-R |
TTGCCTTTGTCCTGTGTGC |
|
11 |
D1y10/D1y12
|
MHDy-F |
GCCCAAGGGCGGATCCTTCT |
68 |
439/403 |
Lee et al. (2025)
|
|
MHDy-/R |
TTGCCCTTGTTCTGGTTGTT |
References
- 1. Ahmad M. 2000. Molecular marker-assisted selection of HMW glutenin alleles related to wheat bread quality by PCR generated DNA markers. Theor Appl Genet 101: 892-896.
- 2. Anderson OD, Greene FC, Yip RE, Halford NG, Shewry PR, Malpica-Romero JM. 1989. Nucleotide sequences of the two high-molecular-weight glutenin genes from the D-genome of a hexaploid bread wheat, Triticum aestivum L. cv Cheyenne. Nucleic Acids Res 17: 461
- 3. Anderson OD, Greene FC. 1989. The characterization and comparative analysis of high-molecular-weight glutenin genes from genomes A and B of a hexaploid bread wheat. Theor Appl Genet 77: 689-700.
- 4. Andrade F, Alvarez JB, Guzman C. 2024. Development of a germplasm master set covering variability of high molecular weight glutenin subunits for the GLU-A1 locus in Triticum sp. J Cereal Sci 118: 103968.
- 5. Boden SA, McIntosh RA, Uauy C, Krattinger SG, Dubcovsky J, Rogers WJ, Xia XC, Badaeva ED, Bentley AR, Brown-Guedira G, Caccamo M, Cattivelli L, Chhuneja P, Cockram J, Contreras-Moreira B, Dreisigacker S, Edwards D, Gonzales FG, Guzman C, Ikeda TM, Karsai I, Nasuda S, Pozniak C, Prins R, Sen TZ, Silva P, Simkova H, Zhang Y. Wheat Initiative2023. Updated guidelines for gene nomenclature in wheat. Theor Appl Genet 136: 72
- 6. Brankovic G, Dodig D, Pajic V, Kandic V, Knezevic D, Djuric N, Zivanovic T. 2018. Genetic parameters of Triticum aestivum and Triticum durum for technological quality properties in Serbia. Zemdirbyste 105: 39-48.
- 7. Butow BJ, Ma W, Gale KR, Cornish GB, Rampling L, Larroque O, Morell MK, Bekes F. 2003. Molecular discrimination of Bx7 alleles demonstrates that a highly expressed high molecular-weight glutenin allele has a major impact on wheat flour dough strength. Theor Appl Genet 107: 1524-1532.
- 8. Choi R, Yu JH, Hong SM, Kim KM, Jung HY, Mo Y, Park CS. 2022. Identification of Korean wheat cultivars using multiplex STS-SSR markers. Korean J Breed Sci 54: 119-129.
- 9. D'ovidio R, Anderson OD. 1994. PCR analysis to distinguish between alleles of a member of a multigene family correlated with wheat bread-making quality. Theor Appl Genet 88: 759-763.
- 10. D'ovidio R, Masci S, Porceddu E. 1995. Development of a set of oligonucleotide primers specific for genes at the Glu-1 complex loci of wheat. Theor Appl Genet 91: 189-194.
- 11. D'ovidio R, Porceddu E, Lafiandra D. 1994. PCR analysis of genes encoding allelic variants of high-molecular-weight glutenin subunits at the Glu-D1 locus. Theor Appl Genet 88: 175-180.
- 12. Gianibelli MC, Gupta RB, Lafiandra D, Margiotta B, MacRitchie F. 2001. Polymorphism of high MrGlutenin subunits in Triticum tauschii: Characterisation by chromatography and electrophoretic methods. J Cereal Sci 33: 39-52.
- 13. Halford NG, Forde J, Anderson OD, Greene FC, Shewry PR. 1987. The nucleotide and deduced amino acid sequences of an HMW glutenin subunit gene from chromosome 1B of bread wheat (Triticum aestivum L.) and comparison with those of genes from chromosomes 1A and 1D. Theor Appl Genet 75: 117-126.
- 14. Heo H, Sherman J. 2013. Identification of QTL for grain protein content and grain hardness from winter wheat for genetic improvement of spring wheat. Plant Breed Biotech 1: 347-353.
- 15. Jang YR, Beom HR, Altenbach SB, Lee MK, Lim SH, Lee JY. 2017. Improved method for reliable HMW-GS identification by RP-HPLC and SDS-PAGE in common wheat cultivars. Molecules 22: 1055
- 16. Jang YR, Cho K, Kim SW, Altenbach SB, Lim SH, Sim JR, Lee JY. 2020. Development of an optimized MALDI-TOF-MS method for high-throughput identification of high-molecular-weight glutenin subunits in wheat. Molecules 25: 4347
- 17. Lafiandra D, Tucci GF, Pavoni A, Turchetta T, Margiotta B. 1997. PCR analysis of x-and y-type genes present at the complex Glu-A1 locus in durum and bread wheat. Theor Appl Genet 94: 235-240.
- 18. Lee JY, Park CS, Kim HJ, Kim JH, Kim MS, Kim YT, Kang CS, Lim SH, Ha SH, Ahn SN, Kim YM. 2013. Two-dimensional electrophoresis of high molecular weight glutenin subunits in Korean wheat cultivars. Korean J Breed Sci 45: 240-252.
- 19. Lee MH, Choi C, Kim KH, Son JH, Lee GE, Choi JY, Kang CS, Sohn J, Ko JM, Kim KM. 2023. Generation of wheat near-isogenic lines overexpressing 1Bx7 glutenin with increased protein contents and SDS-sedimentation values. Plants 12: 1244
- 20. Lee MH, Choi C, Kim KH, Son JH, Park J, Lee GE, Choi JY, Kang CS, Shon J, Ko JM, Kim KM. 2022. Analysis of protein properties and gluten protein composition evaluation of wheat genetic resources. Korean J Breed Sci 54: 245-259.
- 21. Lee MH, Kang CS, Choi MG, Choi JS, Kim MJ, Yoon M, Kim KM, Choi C. 2025. Development of PCR-based markers for the identification of wheat HMW glutenin alleles at GLU-A1 and GLU-D1 loci. Sci Rep 15: 32008
- 22. Lee MH, Kim KY, Kang CS, Yoon M, Jang KC, Choi C. 2024a. Development of PCR-based markers for the identification of wheat HMW glutenin alleles at Glu-1Bx and Glu-1By loci. BMC Plant Biol 24: 395
- 23. Lee MH, Kim KM, Choi MG, Kang CS, Yoon M, Jang KC, Choi C. 2024b. Assessment of allele composition of HMW-GS Glu-B1 in domestic wheat cultivars using PCR-based markers. Korean J Breed Sci 56: 257-268.
- 24. Lee YG, Choi SC, Kang Y, Kang CS, Kim C. 2018. Genomics approach to identify the cause of the missing omega-5 gliadin protein in O-free wheat. Plant Breed Biotech 6: 413-425.
- 25. Lei ZS, Gale KR, He ZH, Gianibelli C, Larroque O, Xia XC, Butow BJ, Ma W. 2006. Y-type gene specific markers for enhanced discrimination of high molecular weight glutenin alleles at the Glu-B1 locus in hexaploid wheat. J Cereal Sci 43: 94-101.
- 26. Liu S, Chao S, Anderson JA. 2008. New DNA markers for high molecular weight glutenin subunits in wheat. Theor Appl Genet 118: 177-183.
- 27. Ma W, Zhang W, Gallle K. 2003. Multiplex-PCR typing of high molecular weight glutenin alleles in wheat. Euphytica 134: 51-60.
- 28. McIntosh RA, Dubcovsky J, Rogers WJ, Xia XC, Raupp WJ. 2020. Catalogue of gene symbols for wheat: 2020 Supplement.
- 29. McIntosh RA. 2024. Catalogue of gene symbols for wheat: 2024 edition (WORD extension of the MACGEN).
- 30. Park CS, Kang CS, Jeung JU, Woo SH. 2011. Influence of allelic variations in glutenin on the quality of pan bread and white salted noodles made from Korean wheat cultivars. Euphytica 180: 235-250.
- 31. Payne PI, Lawrence GJ. 1983. Catalogue of alleles for the complex gene loci, Glu-A1, Glu-B1, and Glu-D1 which code for high-molecular-weight subunits of glutenin in hexaploid wheat. Cereal Res Commun 11: 29-35.
- 32. Ragupathy R, Naeem HA, Reimer E, Lukow OM, Sapirstein HD, Cloutier S. 2008. Evolutionary origin of the segmental duplication encompassing the wheat Glu-B1 locus encoding the overexpressed Bx7 (Bx7OE) high molecular weight glutenin subunit. Theor Appl Genet 116: 283-296.
- 33. Rural Development Administration (RDA).2012. Agricultural science and technology of analysis based on research (Ⅰ). pp. 315-374..
- 34. Schwarz G, Felsenstein FG, Wenzel G. 2004. Development and validation of a PCR-based marker assay for negative selection of the HMW Glu-B1-1d (Bx-6) in wheat. Theor Appl Genet 109: 1064-1069.
- 35. Shewry PR, Halford NG, Tatham AS. 1992. High molecular weight subunits of wheat glutenin. J Cereal Sci 15: 105-120.
- 36. Shin S, Kang CS, Kim KH, Park CS. 2012. Analysis of glutenin compositions in Korean wheat cultivar using SDS-PAGE and PCR. Korean J Breed Sci 44: 245-257.
- 37. Skerritt JH. 1998. Gluten proteins: Genetics, structure and dough quality - A review. AgBiotech 10: 247N-270N.
- 38. Smith RL, Schweder ME, Barnett RD. 1994. Identification of glutenin alleles in wheat and triticale using PCR-generated DNA markers. Crop Sci 34: 1373-1378.
- 39. Thompson RD, Bartels D, Harberd NP, Flavell RB. 1983. Characterization of the multigene family coding for HMW glutenin subunits in wheat using cDNA clones. Theor Appl Genet 67: 87-96.
- 40. Xu Q, Xu J, Liu CL, Chang C, Wang CP, You MS, Li BY, Liu GT. 2008. PCR-based markers for identification of HMW-GS loci in common wheat. J Cereal Sci 47: 394-398.
- 41. Zhang Q, Dong YM, An XL, Wang A, Zhang YZ, Li XH, Gao LY, Xia XC, He ZH, Yan YM. 2008. Characterization of HMW glutenin subunits in common wheat and related species by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-MS). J Cereal Sci 47: 252-261.