적요
To improve resistance to planthoppers and viral diseases, a new rice cultivar named ‘Drimi9ho’ was established. The F1 population, derived from a cross between ‘Cheongcheong’ and ‘Nagdong’, was subjected to anther culture. Subsequent generations were advanced in the field under high selection pressure for agronomic traits and disease resistance. Ultimately, the line ‘CNDH-AC63-2-2-2-1’ was selected through the pedigree method and was named ‘Drimi9ho’ after undergoing yield trials and local adaptability tests. ‘Drimi9ho’ has a heading date of August 13, which corresponds to 107 days after sowing, making it a mid-late maturing variety that heads 3 d later than ‘Cheongcheong’. The culm length of ‘Drimi9ho’ is 57.3 cm, which is 15 cm shorter than that of ‘Cheongcheong’. This reduced culm length contributes to improved tolerance to field lodging. In addition, ‘Drimi9ho’ exhibits acquired resistance to rice black-streaked dwarf virus through introgression from ‘Nagdong’. Compared to ‘Cheongcheong’, ‘Drimi9ho’ shows lower protein content and higher amylose content, which resulted in a higher score for cooked-rice palatability. ‘Drimi9ho’ shows improved milling performance compared to ‘Cheongcheong’. The 1,000-grain weight of ‘Drimi9ho’ is 22.12±0.8 g, which is heavier than that of ‘Cheongcheong’ (19.42±0.8 g). The milled rice yield of ‘Drimi9ho’ is 546.6±3.8 kg/10a, representing a 4.7% increase compared to ‘Cheongcheong’ (522.2±5.8 kg/10a). However, ‘Drimi9ho’ is resistant only to bacterial leaf blight race K1; thus, timely disease management is required to control other races such as K2, K3, and K3a (Registration No. 10610).
서언
벼(
Oryza sativa L.)는 세계 주요 식량 작물 중 하나이며, 전 세계 인구 60% 이상의 칼로리를 책임지고 있다(
Fukagawa & Ziska 2019). 벼는 우리나라 농민 대부분의 생계를 담당하고 있으며, 이에 따라 우리나라의 농업경제활동에도 막대한 영향력을 미치고 있다(
Jeong et al. 2021). 또한 벼는 세계 식량 안보에도 중요하게 작용하고 있다(
Bandumula 2018). 지구온난화에 따라 전 세계 기온은 상승하고 있으며, 이는 벼 수량 감소 및 멸구류를 포함한 벼 주요 가해 해충들의 대발생을 가능하게 한다(
Seo et al. 2019,
Yuan 2012). 기온 상승에 따라 흰등멸구(white-backed planthopper,
Sogatella furcifera)의 발생 증가는 벼 생육단계에 따라 서로 다른 반응이 나타나며, 최종적으로 수량 및 미질에 심각한 부정적인 영향을 미친다(
Zhou et al. 2013).
멸구류는 열대 및 아열대 기후 지역에서 생활하며 번식한다(
Kim & Park 2018). 그렇기 때문에 사계절이 뚜렷한 우리나라에서는 월동이 불가능 하며, 매년 6월 중순부터 중국 남부 또는 베트남에서 비래하여 한국과 일본과 같은 동아시아 온대기후의 지역에서 증식한다(
Shin et al. 2021). 멸구류는 벼 줄기 하단부에 밀집하여 체관부의 수액을 흡즙하여 벼를 빠르게 마르게 하고, 번식속도가 빨라 벼 포장의 넓은 범위의 벼가 갈색으로 변하는 집중고사현상인 호포번(hopper burn)이 나타나게 된다(
Adiredjo & Kiswanto 2025). 특히 흰등멸구는 벼에 직접적인 피해를 주기도 하지만 벼남방검은줄오갈병(southern rice black-streaked dwarf virus, SRBSDV)와 같은 바이러스를 매개하여 간접적인 피해를 야기한다(
Zhou et al. 2013). 벼남방검은줄오갈병에 의해 베트남과 중국에서는 30-50%의 벼 수량감소가 발생하였으며, 이는 벼의 정상적인 성장을 방해하여 수량 감소에 따른 농가의 경제적 손실에 직접적인 영향을 준다(
Alonso et al. 2019,
Hoang et al. 2011).
농가에서는 흰등멸구와 같은 해충을 방제하기 위해 살충제 처리와 같은 화학적 방제에 의존하고 있다(
Reddy et al. 2022). 화학적 방제는 단기적으로 병해충 발생을 억제할 수 있다. 하지만 흰등멸구는 환경변화에 대한 적응이 빠르며, 이는 살충제에 대한 저항성 변이가 빨리 발생하며 내성이 생긴 개체에 의해 저항성 붕괴가 빠르게 일어나는 요인으로 작용할 수 있다(
Yangon 2011). 이미 화학적 방제의 남용으로 인해 멸구류에서 유기인계, 페닐피라졸계 등과 같은 살충제 성분에 대한 다양한 저항성 발달이 보고되고 있다(
Horgan et al. 2020). 화학적 살충제는 이외에도 토양 및 수질과 같은 환경오염에 대한 문제도 야기할 수 있다(
Ali et al. 2020). 하지만 저항성 품종을 육성 할 경우 화학적 방제를 하는 것보다 병해충 저항성의 지속력이 장기적이며, 친환경적인 방법이다(
Kim et al. 2019).
최근 우리나라의 기온 상승에 따른 각종 기상재해들의 발생 빈도는 증가 하고 있으며, 병해충의 발생 양상도 다양화 되고 있다. 이러한 현상은 벼 수량 감소에 직접적인 영향이 되고 있으며, 이에 대한 대비가 필요하다. 시대가 변화함에 따라 우리나라 국민들은 건강에 대한 기대감은 증가하고 있다. 이러한 요인들에 대응하기 위해 경북대학교 응용생명과학과 식물분자육종학 연구실에서는 흰등멸구에 대한 저항성이 향상된 품종인 ‘드리미9호’를 육성하였다. ‘드리미9호’에 대한 주요 농업적 특성 및 육성 경위를 함께 제시하였으며, 변화하고 있는 기후와 소비자들의 인식에 대한 맞춤형 벼 품종 육성에 활용 하고자 한다. 이에 따라 ‘드리미9호’의 육성 경위와 주요 특성을 보고하는 바이다.
재료 및 방법
시험재료 및 재배방법
‘드리미9호’를 육성하기 위한 교배 모본으로 ‘청청’, 부본으로는 ‘낙동’을 선정하였다. ‘청청’과 ‘낙동’은 경북대학교 농업생명과학대학 부속실험실습장에서 교배하였으며, 인공교배를 통한 F1 종자 수확 및 식물체를 양성하였다. 이후 약배양을 통해 배가반수체 집단을 육성하였으며, 매년 포장에서 세대진전 시켜 120 계통으로 구성된 배가반수체 집단을 유지하였다. 모든 종자는 파종 전 스포탁(spotak, HANKOOKSAMGONG, Seoul, South Korea)을 이용하여 종자 소독하였으며, 25℃에서 4일간 침종 및 최아 시켰다. 소독된 종자는 경북대학교 농업생명과학대학 부속실험실습장에서 파종하였다. 파종량은 포장으로 이앙하는데 필요한 식물체의 수보다 10% 많은 양을 파종하였다. 파종 후 30일이 지났을 때 포장으로 이앙하였으며, 보통기 재배를 하였다. 포장에서 재식 거리는 30×15 cm로 하였다. 각 계통당 6줄씩 이앙하였으며, 각 줄당 25개의 식물체가 이앙되도록 하였다. 계통유지 및 세대진전을 위해 주당 1본씩 손 이앙하였으며, 생산력 검정 시험을 수행 할 때는 주당 3본씩 구당 150주를 이앙하였으며, 3반복으로 보통기 재배하였다. 벼 생육 기간 동안의 시비량은 농촌진흥청에서 제시하고 있는 농업과학기술 연구 조사 기준을 따랐으며, N-P2O5-K2O= 9-4.5-5.7 kg/10a로 시비하였다. 질소 비료는 시비-분얼비-수비를 각각 50-20-30의 비율로 분시하였으며, 인산은 전량 기비로 하였다. 칼륨은 기비-수비를 70-30으로 분시하였다. 시비량 이외의 벼 생육 기간동안 필요한 재배 관리 방법들은 모두 농촌진흥청에서 제시하고 있는 표준재배방법을 준수하였다(RDA 2017). 벼 주요 농업 형질과 생육 특성, 수량, 생리 장해 및 병해충 저항성, 도정특성과 관련된 조사 방법들은 모두 농촌진흥청에서 제시하고 있는 신품종개발 공동연구사업 과제 수행계획서와 농업과학 기술 연구조사 분석기준에서 제시된 방법에 의하여 조사되었다(RDA 2018).
주요 농업 형질 조사
경북대학교 농업생명과학대학 부속실험실습장의 포장에서 각 시험 재료들의 포장 및 낟알 특성과 관련된 농업형질들을 조사하였다. 동일한 계통에서 출수가 약 50% 이루어진 시점에 출수기를 조하였다. 출수기 이외의 농업 형질들은 포장에서 계통들이 성숙기가 되었을 때 조사하였다. 각 계통에서 무작위로 20개체를 선발하여 간장, 수장, 수수를 조사하였다. 지면에서 이삭목까지의 길이를 간장으로 조사하였으며, 이삭목부터 이삭의 가장 긴 부분까지의 길이를 수장으로 조사하였다. 하나의 식물체에서 발생한 모든 수수의 개수를 수수로 조사하였다. 그리고 출수 후 50일이 지났을 때, 각 계통에서 무작위로 5주씩 예취하여 수당립수, 임실률, 등숙률을 조사하였으며, 모든 조사는 독립적인 3반복 실험으로 진행하였다. 수량조사현미기를 이용하여 수확한 정조 1 kg을 제영하여 정현비율을 조사하였으며, 정조 수량에 정현비율을 곱하여 현미수량을 계산 후 10 a당 수량으로 환산하였다. 백미 수량을 계산하기 위해 일반적으로 통용되는 현백률인 0.92를 현미수량에 곱하여 계산하였으며, 제현된 현미를 이용하여 천립중을 측정하였다(
Park et al. 2020).
낟알 특성을 조사하기 위해 각 공시 재료의 현미를 무작위로 30립을 선발하였으며, 버니어캘리퍼스(Caliper CD-15CP, Mitutoyo Corp., Japan)를 이용하여 낟알 길이, 너비, 폭을 조사하였다. 현미 장폭비는 낟알 너비에 대한 낟알 길이의 비율로 계산하였다. 백미의 품질에 대한 평가를 위해 백미의 투명도, 심복백 정도, 알칼리붕괴도, 아밀로스 및 단백질 함량을 조사하였다. 백미의 투명도를 조사하기 위해 grain polisher (pearlest, cat. TP-3000, kett, Tokyo, Japan)를 이용하여 현미를 백미로 만들었으며, 이후 백미에 대한 투명도를 달관 조사하였다. 백미의 투명도를 달관조사 하기 위해 백미 30립을 무작위로 조사하였으며, 1-9점을 투명도에 따라 부여하였다. 백미의 투명도가 유리알처럼 매우 맑을 경우에는 1점을 부여하였으며, 백미의 투명도가 중간일 때는 5점, 매우 불투명할 경우에는 9점을 부여하였다. 백미의 심복백도 달관 조사하여 정도에 따라 0-9점을 부여하였다. 심복백이 전혀 없을 경우에는 0점, 전체 면적의 5% 이하가 심복백일 경우 1점, 6-10%일 경우 3점, 11-20%일 경우 5점, 21-40%일 경우 7점, 41% 이상일 경우 9점을 부여하였다. 알칼리 붕괴도 조사는 90×15 mm의 페트리디쉬(petridish, cat. 10090, SPL, Gyeonggi-do, Korea)에서 백미를 1.4% KOH 용액과 반응시켜 조사하였다. 페트리디쉬에 백미 6립을 일정한 간격으로 배치하고 1.4% KOH 용액을 10 mL을 넣어 주었으며, 30℃의 항온기에서 23시간 정치 후 백미의 퍼짐 정도와 투명도를 달관조사하여 1-7점을 부여하였다. 1.4% KOH 용액과 반응하였을 때 백미가 반응 전과 변함없이 부풀지 않았을 때 1점, 반응전과 비슷한 상태를 유지하고 있지만 백미가 약간 부풀었을 때 2점, 백미가 부푼 것이 확연히 느껴지고 백미의 외관에 금이 발생하기 시작하여 약간 퍼진 경우에는 3점, 백미의 표면에 금이 발생하여 갈라진 것이 확연히 보이며, 백미 주변으로 퍼짐 현상이 보이지만 투명화가 진행되지 않았을 때는 4점, 백미가 심하게 갈라지고 투명화가 시작되었을 때는 5점, 백미가 완전히 퍼지고 백미의 외곽이 완전히 투명화 되었을 때는 6점, 투명화가 완벽히 진행되어 백미의 형체를 완전히 알아 볼 수 없을 때는 7점을 부여하였다. Near-infared spectroscopy (Kett, AN-820, Japoan)을 이용하여 공시재료들의 아밀로스 및 단백질 함량을 분석하였다(
Chemists & Horwitz 1960).
도열병 저항성 검정을 위해 7월 초순에 시험재료를 10×20 cm 간격으로 밭못자리에 파종하였다. 파종 후 시비는 질소-인산-가리를 24-9-9 kg/10a로 하였다. 공시 품종 주변에는 도열병에 대한 이병성 품종인 ‘호평’을 spreader로 함께 파종하였다. 도열병 저항성 검정은 파종 후 30일이 지났을 때 하였으며, IRTP (International Rice Testing Program)에서 제시하고 있는 밭못자리 도열병 표준 검정법에 근거하여 조사하였다(0-3점: 저항성, 4-6점: 중도저항성, 7-9점: 감수성,
Rice 1996).
벼 흰잎마름병 저항성 검정을 위해 농촌진흥청 국립식량과학원 작물환경과에서 흰잎마름병 균주인 HB01013 (K
1), HB01014 (K
2), HB01015 (K
3), HB01009 (K
3a)를 분양 받았다. 분양받은 균주를 활용하여 검정 계통이 최고분얼기가 되었을 때 각 균주별로 식물체의 엽선단 5 cm 부위에 가위절엽법으로 접종하였다. 저항성 검정은 균주 접종 후 3주가 지났을 때 접종한 개체에서 병반이 가장 긴 3개 잎의 병반길이를 측정한 값을 평균 계산하였다. 5 cm 이하는 저항성, 5-10 cm는 중도 저항성, 10 cm 이상은 이병성으로 질적 저항성을 판정하였다(
Park et al. 2016).
멸구류에 대한 저항성 검정은 생물 검정을 통해 확인하였다. 저항성 검정을 위해 사용된 멸구류는 농촌진흥청 국립식량과학원 작물환경과에서 분양 받았다. 분양 받은 멸구류 암수 50쌍은 경북대학교 온실에서 사육하였다. 사육 케이지는 투명 아크릴판으로 제작하여 빛이 투과할 수 있도록 하였으며, 가로 50 cm, 세로 50 cm, 높이 40 cm가 되도록 하였다. 사육 케이지의 옆면에는 직사각형 모양의 가로 40 cm, 높이 30 cm의 100 mesh망으로 3면을 만들어 공기의 순환이 원활하게 될 수 있도록 하였다. 사육 케이지의 앞면에는 먹이 공급을 효과적으로 하기 위해 가로 40 cm, 세로 30 cm의 미닫이 문을 설치하였다. 멸구류를 사육하기 위해 ‘추청’을 식이재료로 사용하였다. ‘추청’ 종자를 소독 하여 파종 후 3-4엽기가 되었을 때 식이재료로 공급하였으며, 7일마다 새로운 먹이로 교환 해주었다.
공시 재료들의 멸구류에 대한 저항성 검정을 위해 ‘청청’을 저항성 품종, ‘추청’을 감수성 품종으로 공시하였다. 시험재료들은 종자를 소독 후 가로 32 cm, 세로 23 cm, 높이 10 cm의 플라스틱 상자에 시험 재료당 20립씩 2열로 핀셋 파종하였으며, 3-4엽기가 되었을 때 멸구류를 접종 시켜 저항성 검정을 하였다. 각 계통의 간격은 2 cm로 하였으며, 저항성 검정은 독립적인 3반복으로 실시하였다. 멸구류의 저항성 검정을 위해 2-3령충을 각각의 유묘마다 10-15마리가 접종 될 수 있도록 하였다. 시험재료의 저항성 검증은 감수성 품종인 ‘추청’이 완전히 고사하였을 때를 기점으로 각 계통들의 차이를 벼 표준평가방법에 준하여 저항성과 이병성으로 판정하였다(
Rice 1996).
검은줄오갈병, 오갈병, 줄무늬잎마름병에 대한 ‘드리미9호’의 유묘저항성을 평가하기 위해 가로 32 cm, 세로 23 cm, 높이 10 cm의 플라스틱 상자에 소독 후 최아된 종자를 20립씩 1열로 핀셋 파종하였다. 시험재료가 2열씩 파종 되었을 때 검은줄오갈병, 오갈병, 줄무늬잎마름병 모두에 대해 저항성 품종인 ‘낙동’과 이병성 품종인 ‘추청’을 대비품종으로 번갈아 파종하였다. 저항성 검정을 위해 시험재료 파종 후 3-4엽기가 되었을 때 바이러스 보독충을 방사하였으며, 유묘당 10-15마리가 접종되도록 하였다. 바이러스병에 대한 저항성 판정은 보독충 접종 후 감수성 대비 품종인 ‘추청’이 완전히 고사하 시점에 하였으며, 저항성 판정 기준은 농업과학기술 연구조사 분석 기준으로 판정하였다(
Kwak et al. 2007).
시험재료들의 농업형질에 대한 통계분석은 R (Version 4.3.1, The R Foundation for Statistical Computing Platform)을 활용하여 수행 되었다. 조사된 농업 형질들의 평균 및 표준편차를 계산하였으며, 육성 품종과 대비품종간의 비교를 위해 t-test를 적용하였다. 모든 실험 결과는 독립적인 3반복 실험을 통해 얻어진 결과이다.
결과 및 고찰
품종 육성 경위
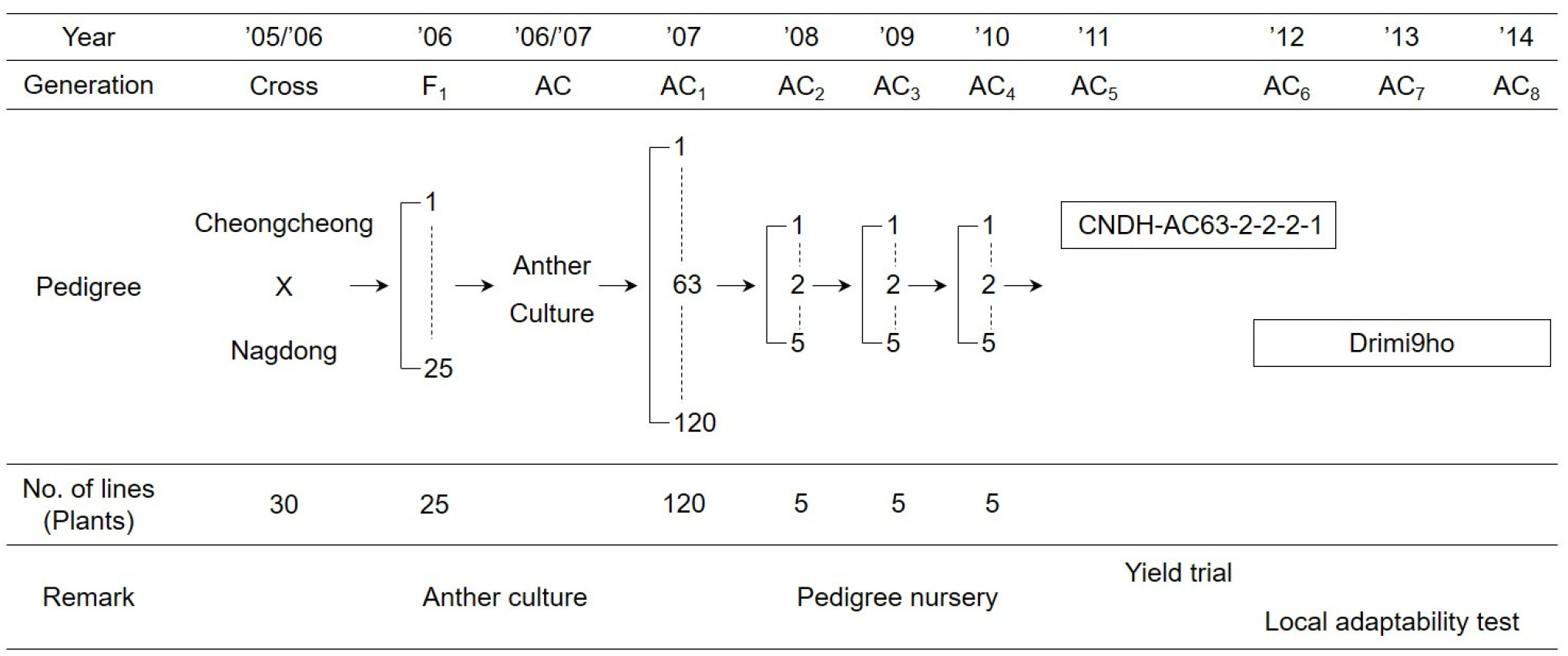
‘드리미9호’는 흰등멸구에 대한 저항성이 향상된 품종을 육성할 목적으로 2005년도에 경북대학교 농업생명과학대학 부속실험실습장에서 흰등멸구에 대한 저항성이면서 다수성인 ‘청청’과 바이러스병에 대한 저항성인 ‘낙동’을 인공교배하여 얻어진 F
1을 약배양 하여 육성하였다. 2006년에는 F
1 식물체가 출수 직전인 상태(엽이간장이 3 cm 나왔을 때)에서 약을 채취하여 배양하였으며, 약배양으로 조기에 고정된 배가반수체집단을 육성하였다. 확보된 약배양 계통을 포장에 전개하여 포장 특성이 우수하고 흰등멸구에 대한 저항성이 있는 우량계통 ‘CNDH-AC63-2-2-2-1’을 선발하여 2011~2012년에 생산력검정시험을 하였으며, 최종적으로 ‘드리미9호’로 명명하였다(
Figs. 1 and
2). 2012~2014년의 3년 동안 보통기 보비조건에서 지역적응시험을 수행하였으며, 표준품종은 ‘청청’을 사용하였다.
Fig. 3은 시험포장에서 ‘드리미9호’가 출수 후 30일이 지난 황숙기의 생육 모습이다. ‘드리미9호’를 보통기 보비재배 하였을 때 파종 후 107일이 된 시점인 8월 13일에 출수하였으며, 청청의 출수기(8월 10일)보다 3일 느린 중만생종이다(
Table 1). ‘드리미9호’는 ‘청청’보다 황숙기에 진한 녹색을 나타내며, ‘청청’보다 초장이 짧고 직립성이 우수하며 초형이 양호하다. ‘드리미9호’의 간장은 57.3±3.9이며, ‘청청(72.3±4.9)’보다 15 cm짧은 단간이다. 수장은 ‘청청(20.5±1.6 cm)’보다 3 cm 짧은 17.4±4.0 cm이며, 수수는 12.3±3.5개로 ‘청청(9.3±4.4)’보다 3개 많은 수준이었다. ‘드리미9호’의 수당립수는 112±3.4개, 등숙비율은 93.74±2.8%이며, ‘청청’의 수당립수와 등숙비율은 각각 108±4.2개, 91.5±1.5%이다. ‘드리미9호’는 ‘청청’보다 수당립수는 3.4±1.5개 많으며, 등숙비율도 2.4±1.2% 높은 수준이었다. ‘드리미9호’의 천립중은 22.12±0.8 g이며, ‘청청(19.42±0.8 g)’보다 2.4±0.4 g 더 무거웠다.
생리장해 저항성
‘드리미9호’의 생리장해 특성을 ‘청청’과 비교하기 위해 못자리에서 50일간 생육 시킨 후 포장으로 이앙하였다. ‘드리미9호’를 포장으로 이앙하였을 때 불시출수와 위조현상은 모두 관찰되지 않았다(
Table 2). ‘드리미9호’는 ‘청청’보다 하엽의 노화 속도가 느린 편이었다. 내냉성 검정하였을 때 ‘드리미9호’와 ‘청청’의 유묘 적기는 비슷한 수준이었다. 또한 냉해로 인해 ‘드리미9호’는 8±1.3일간 출수가 지연 되었지만, ‘청청’은 14±1.5일 지연되어 ‘청청’보다 출수지연 일수는 6±1.2일 짧았다. 또한 냉해조건에서 ‘드리미9호’의 임실율은 35.3±2.5%이며, ‘청청(21.8±1.8%)’보다 통계적으로 유의미한 수준으로 임실율이 높게 조사되었다. 저온 발아율은 ‘드리미9호’와 ‘청청’이 각각 79.5±1.8%, 80.1±1.6%로 통계적으로 동일한 수준이었다. ‘드리미9호’의 수발아율은 42.4±3.2%이며, ‘청청(51.4±3.8%)’보다 낮았다. ‘드리미9호’는 ‘청청’에 비해 냉해 저항성이 비슷하거나 강화 되었다.
포장에서 ‘드리미9호’의 도복 저항성 정도를 분석하기 위해 초장, 3절 간장, 좌절중, 도복지수, 포장에서의 도복 발생 정도를 ‘청청’과 비교분석하였다(
Table 3). ‘드리미9호’의 초장은 74.7±5.3 cm로 ‘청청(92.8±3.8 cm)’보다 약 8 cm 짧았다. ‘드리미9호’의 3절 간장과 좌절중은 각각 8.3±1.2 cm, 1,142±3.2 g으로 ‘청청(10.2±0.8 cm)’보다 3절 간장은 2.1±0.9 cm 짧았으며, 좌절중은 ‘청청(994.7±3.4 g)’보다 높았다. 포장에서 ‘드리미9호’의 도복지수는 163±5.3으로 ‘청청(212±4.8)’보다 도복지수가 낮아 도복에 안정적인 특성을 나타냈다. ‘드리미9호’는 ‘청청’보다 초장과 3절 간장이 짧았다. 또한 좌절중은 높고 도복지수는 낮았다. ‘드리미9호’와 ‘청청’은 모두 포장에서 도복이 발생하지 않았지만 도복 저항성과 관련된 특성에서 ‘드리미9호’가 ‘청청’보다 향상 되었다.
‘드리미9호’의 도열병, 흰잎마름병, 바이러스병, 멸구류에 대한 저항성 평가를 하였다. ‘드리미9호’의 잎도열병 이병율을 밭못자리에서 검정하였을 때 저항성 정도는 약 3.6±1.1이었으며, ‘청청(3.0±1.2)과 동일한 수준으로 저항성이 있는 것으로 평가 되었다(
Table 4). ‘드리미9호’의 목도열병 이병수율은 약 2.0±0.2%이며, ‘청청(1.8±0.3%)’과 통계적으로 동일한 수준이었다. ‘드리미9호’는 잎도열병과 목도열병에 대한 저항성 정도가 ‘청청’과 유사한 수준이었다.
흰잎마름병에 대한 저항성 평가를 위해 ‘드리미9호’와 ‘청청’에 벼흰잎마름병 유도 균주 K
1 (HB1013), K
2 (HB1014), K
3 (HB1015), K
3a (HB1009)를 접종하였다. ‘드리미9호’는 ‘청청’과 동일하게 K
1에 대해서는 저항성을 나타냈다. 하지만 K
2, K
3, K
3a에 대해서는 모두 이병성이다(
Table 5).
‘드리미9호’에서 우리나라의 대표적인 바이러스병 줄무늬잎마름병, 오갈병, 그리고 검은줄오갈병에 대한 저항성을 평가하였다(
Table 5). ‘드리미9호’의 줄무늬잎마름병에 대한 이병률은 15.8±2.3%, 오갈병에 대한 이병률은 18.6±2.8%, 검은줄오갈병에 대한 이병률은 45.5±2.4%로 조사되었다. ‘드리미9호’는 줄무늬잎마름병과 오갈병에 대해서는 저항성을 나타냈으며, 검은줄오갈병에 대해서는 중도저항성이 있는 것으로 조사 되었다. ‘청청’은 줄무늬잎마름병과 오갈병에 대한 저항성은 있었지만 검은줄오갈병에 대해서는 이병성이었다. 따라서 ‘드리미9호’는 ‘청청’에서 검은줄오갈병에 대한 저항성이 추가 되었다. 또한 ‘드리미9호’와 ‘청청’은 멸구류에 대해서 모두 저항성이 있는 것으로 조사 되었다.
‘드리미9호’는 잎도열병과 목도열병에 대해서는 ‘청청’과 유사한 수준으로 저항성이 있었다. 또한 ‘드리미9호’는 ‘청청’과 동일하게 우리나라 주요 흰잎마름병 발생 균주인 K1에 대해서만 저항성이었으며, K2, K3, K3a에 대해서는 모두 이병성이었다. 하지만 ‘드리미9호’는 ‘청청’보다 오갈병과 검은줄오갈병에 대한 저항성이 모두 향상 되었다. 또한 ‘드리미9호’와 ‘청청’은 멸구류에 대해 모두 저항성이다. 종합하면 ‘드리미9호’는 ‘청청’과 비슷한 수준의 저항성이지만 바이러스병에 대한 저항성이 강화 되어 우리나라의 주요 병해충에 대한 복합 저항성을 가지는 품종으로 육성 되었다.
미질 및 도정 특성
‘드리미9호’의 현미 낟알 길이, 너비, 장폭비를 조사하여 ‘청청’과 낟알의 특성을 비교분석하였다(
Table 6). ‘드리미9호’의 현미 낟알 길이, 너비, 두께는 각각 5.12±0.8 mm, 2.62±1.2 mm, 1.82±1.3으로, ‘청청(5.08±1.1 mm, 2.65±1.1 mm, 1.76±1.1)’과 낟알 특성이 모두 통계적으로 유의한 차이없이 동일한 수준인 단원형의 단립종 특성을 보였다. ‘드리미9호’의 외관상품위를 달관 조사하였을 때 ‘드리미9호’는 심백과 복백이 없었으며, 현미가 맑고 투명하였다. ‘청청’은 심백과 복백이 모두 있었으며, ‘드리미9호’는 ‘청청’보다 외관품위가 향상 되었다. ‘드리미9호’의 알칼리 붕괴도는 6.2±0.2 수준이며, ‘청청(6.7±0.2)’과 비슷하거나 동일한 수준이었다. ‘드리미9호’의 단백질과 아밀로스 함량은 각각 5.1±1.3%, 18.6±1.5%로 ‘청청(6.9±1.2%, 17.8±1.3%)’보다 단백질 함량은 감소되고 아밀로스 함량은 증가되어 ‘드리미9호’는 ‘청청’보다 미질 특성이 향상 되었다. 또한 식미관능시험을 통해 ‘드리미9호’의 밥맛, 찰기, 밥알의 모양, 질감, 밥을 했을 때의 냄새를 종합적으로 판단하였을 때 ‘드리미9호’는 0.21±0.1의 평가를 받았으며, ‘청청’은 0.12±0.3의 평가를 받아 식미는 ‘드리미9호’가 ‘청청’보다 향상되었다고 평가 받았다.
도정특성을 분석하였을 때 ‘드리미9호’의 제현률과 현백률은 각각 82.5±2.5%, 89.3±3.2%로 ‘청청(81.3±3.4%, 88.4±2.5%)’보다 높아 도정률이 75.3±1.2%로 ‘청청(72.5±3.4%)’보다 우수하였다(
Table 7). ‘드리미9호’와 ‘청청’의 완전미율은 각각 91.6±2.5%, 89.8±2.3%로 동일한 수준이었다. 하지만 ‘드리미9호’의 완전미도정수율은 68.4±3.4%로 ‘청청(64.2±3.5%)’보다 완전미도정수율이 개선되었다고 판단된다.
‘드리미9호’와 ‘청청’의 현미의 낟알 길이, 너비, 폭은 동일한 수준이었으며, 모두 단원형에 단립종이었다. 하지만 ‘청청’은 심백과 복백이 모두 있었지만 ‘드리미9호’는 심복백이 모두 관찰 되지 않았으며, 현미의 외관품위가 향상되었다. 또한 식미를 구성하는 물리화학적 구성 요소인 아밀로스와 단백질 함량을 분석하였을 때 ‘청청’보다 아밀로스 함량은 증가한 반면 단백질 함량이 감소되어 식미가 향상되었을 것으로 기대하였다. 하지만 물리화학적 요소들이 우수하게 분석되었다고 해서 실제 사람이 느끼는 식미는 다르게 평가될 수 있기 때문에 식미관능검정시험을 하였다. ‘드리미9호’는 식미관능검정시험에서도 ‘청청’보다 우수한 성적으로 평가 받았으며, 이는 ‘드리미9호’가 실험적인 요소 이외에도 실제 밥으로 먹었을 때도 느끼는 식미도 우수하였다. 따라서 ‘드리미9호’는 ‘청청’보다 현미외관품위 및 식미 특성이 보강되어 육성 되었다고 판단된다.
수량
‘드리미9호’의 수량을 조사하기 위해 영남지역 3개소인 대구, 군위, 칠곡에서 보통기 재배를 하였다(
Table 8). 영남지역에서 ‘드리미9호’와 ‘청청’을 보통기 재배시험 하였을 때 각각 546.6±3.8 kg/10a, 522.2±5.8 kg/10a였다. 영남지역에서 보통기 재배하였을 때 ‘드리미9호’는 ‘청청’보다 수량이 통계적으로 유의한 수준에서 약 4.7% 증수하였다. ‘드리미9호’는 3년간 대구, 군위, 칠곡에서 지역적응성 시험을 하였을 때 3년간 모두 ‘청청’보다 높은 수량을 나타냈다.
‘드리미9호’는 포장 특성 및 현미외관품위가 ‘청청’보다 우수하다. 또한 우리나라에서 빈번하게 발생하여 벼 수량 및 품질에 직접 또는 간접적인 영향을 미치고 있는 도열병, 흰잎마름병(K1), 줄무늬잎마름병, 오갈병, 검은줄오갈병, 멸구류에 대해서 강한 저항성이다. 이러한 특성이 농민에게 보급된 후 유지되기 위해서 재배상유의점을 철저히 지켜져야한다. 키다리병 방제를 위해서 파종 전 종자소독을 할 때 소독약의 매뉴얼을 준수하여 종자 소독을 철저하게 해야한다. ‘드리미9호’는 단간내도복이기 때문에 재배할 때는 농촌진흥청에서 제시하고 있는 표준 시비방법에 따라 저적 비료를 시비하였을 때 포장 도복 발생이 없으며 밥맛이 우수하게 유지될 수 있다. 질소질 비료를 과다하게 사용하게 되면 포장 도복이 발생하여 등숙 및 품질이 저하될 수 있으며 병해충 발생이 우려되기 때문에 적정 균형 시비가 되어야 한다. ‘드리미9호’는 흰잎마름병 균계 중 K1에 대한 저항성만 있기 때문에 적기 기본 방제를 통한 흰잎마름병 균계(K2, K3, K3a)에 대한 대비가 필요하다. 흰잎마름병 방제를 위해 등록된 약제를 안전사용기준을 준수하여 적기에 철저한 살포가 필요하다.
적요
‘드리미9호’는 경북대학교 식물분자육종학 연구실에서 우리나라 자포니카 벼 품종의 흰등멸구 저항성 향상을 위해 육성 되었다. ‘드리미9호’는 ‘청청’과 ‘낙동’을 단교잡 인공교배 하여 육성 되었다. 조기에 고정계통을 확보하기 위해 F1 식물체를 약배양하였으며, 배가반수체집단을 경북대학교 농업생명과학대학 부속실험실습장에서 세대진전 및 포장특성을 조사하였다. 배가반수체집단의 계통육성 과정 중에서 도복에 강하고 병 저항성이 향상된 우량계통을 선발하였으며, 생산력검정시험과 지역적응성검정시험을 거쳐 ‘드리미9호’가 육성 되었다. ‘드리미9호’는 보통기 보비재배를 하였을 때 평균 출수기가 8월 13일로 ‘청청’보다 3일 늦은 중만생종이다. ‘드리미9호’의 간장은 57.3 cm로 ‘청청’보다 15 cm 짧은 단간이며 좌절중이 높아 포장에서 도복에 강한 특성을 나타냈다. ‘드리미9호’의 현미 천립중은 22 g으로 ‘청청’에 비해 무거웠으며 수량은 영남지역에서 546.6±3.8 kg/10a로 ‘청청’보다 4.7% 증수하였다. 또한 ‘드리미9호’는 ‘청청’보다 현미외관품위가 향상되었으며 아밀로스함량은 증가되고 단백질 함량은 감소되어 우수한 식미 특성을 가졌다. ‘드리미9호’는 도열병, 줄무늬잎마름병, 오갈병, 검은줄오갈병, 멸구류에 대해서는 강한 저항성을 나타낸다. 하지만 흰잎마름병에 대해서는 오직 K1 균계에만 저항성을 나타내기 때문에 흰잎마름병 발생을 대비하기 위한 적기 방제가 필요하다. ‘드리미9호’는 포장특성 및 재배안정성이 우수하며, 외관품위가 좋다. 또한 흰등멸구에 대한 저항성이 향상되어 기후 변화에 따른 국내 자포니카 품종의 재배안정성의 향상에 기여할 수 있는 품종으로 육성 되었다.
사사
This work was carried out with the support of “Cooperative Research Program for Agriculture Science and Technology Development (Project No. RS-2024-00322408)” Rural Development Administration, Republic of Korea.
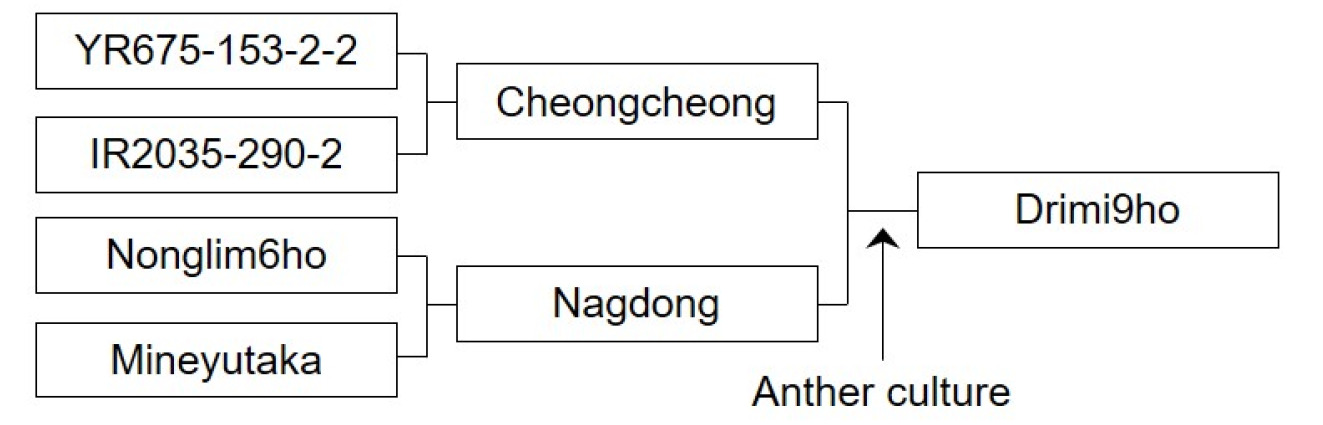
Fig. 1Genealogical and schematic diagram of the breeding process for ‘Drimi9ho’. ‘Drimi9ho’ was developed to improve resistance to both insect pests and viruses. F1 plants derived from a cross between ‘Cheongcheong’ and ‘Nagdong’ were subjected to anther culture. The pedigree breeding method was applied. During field generations, plants were evaluated for agronomic traits and resistance to planthopper. The line ‘CNDH-AC63-2-2-2-1’ was selected and subsequently evaluated through observation yield trials, replicated yield trials, and local adaptability tests, after which it was finally named ‘Drimi9ho’.
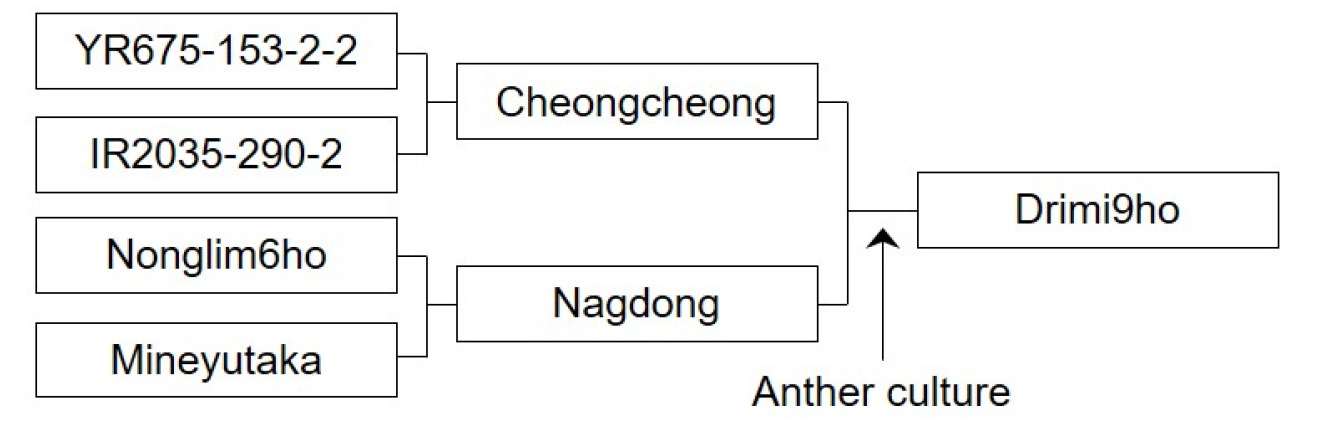
Fig. 2Pedigree diagram of ‘Drimi9ho’. ‘Drimi9ho’ was developed by crossing ‘Cheongcheong’, which confers resistance to planthopper, with ‘Nagdong’, which has resistance to viral diseases. The derived F1 plants were subjected to anther culture, and selection was conducted using the pedigree breeding method based on comprehensive evaluation of field performance and grain appearance. The final selected line was designated as ‘Drimi9ho’.
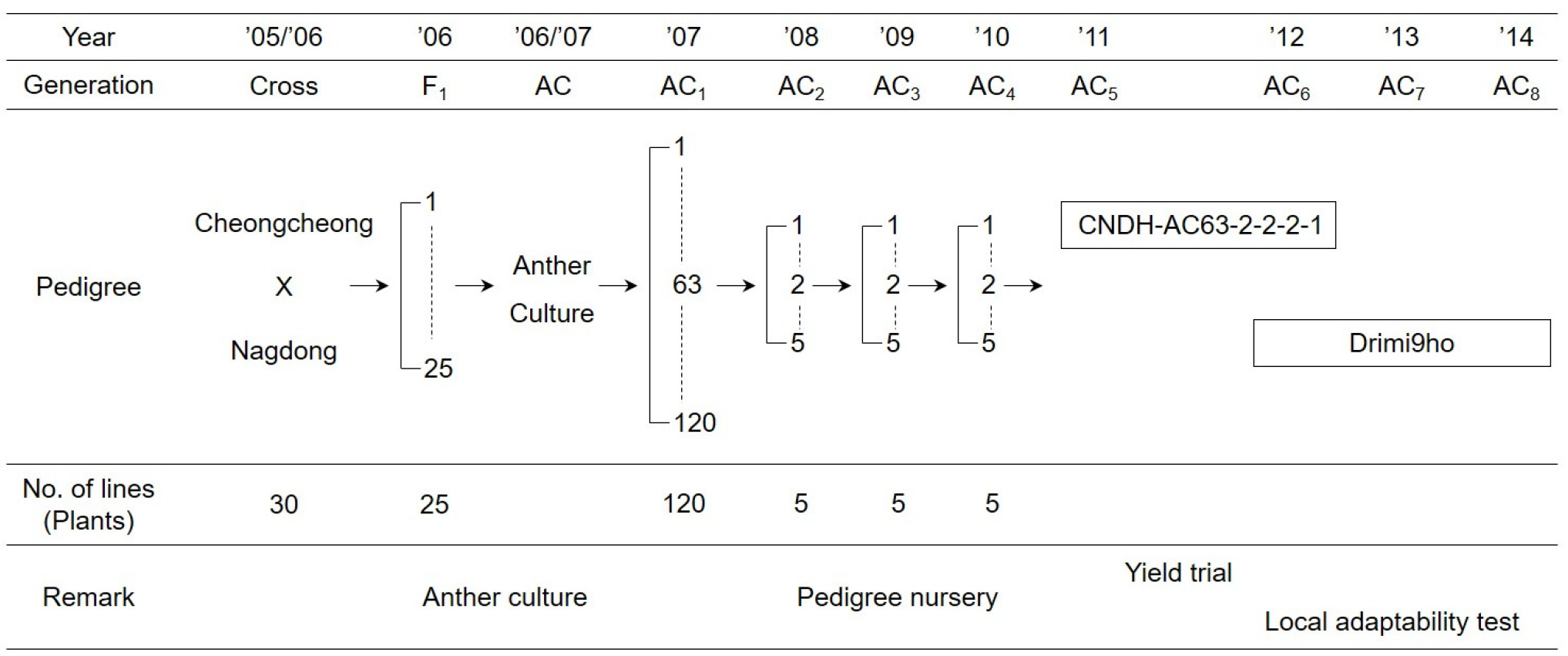
Fig. 3Phenotype of ‘Drimi9ho’ at the ripening stage in the field. Compared to ‘Cheongcheong’, ‘Drimi9ho’ has a 15 cm shorter culm length and a 3 cm shorter panicle length. However, ‘Drimi9ho’ has approximately three more panicles per hill and has a greater number of spikelets per panicle. In addition, the 1,000-grain weight of ‘Drimi9ho’ is higher than that of ‘Cheongcheong’. These traits contribute to improved lodging tolerance and enable higher yield performance in the field. Scale bar: 10 cm.

Table 1Field performance and yield-related agricultural traits of ‘Drimi9ho’.
Table 1
|
Cultivar |
Heading
date
(DAS)z
|
Culm
length
(cm) |
Panicle
length
(cm) |
No. of
panicles
per hill |
No. of
s
pikelets
per panicle |
Ratio of ripened
grain (%) |
1,000-grain weight of brown rice (g) |
|
Drimi9ho |
107±0.9** y
|
57.3±3.9**
|
17.4±4.0**
|
12.3±3.5**
|
112±3.4**
|
93.74±2.8**
|
22.12±0.8**
|
|
Cheongcheong |
104±1.3 |
72.3±4.9 |
20.5±1.6 |
9.3±4.4 |
108±4.2 |
91.5±1.5 |
19.42±0.8 |
Table 2Physiological adaptation of ‘Drimi9ho’ to different abiotic stresses.
Table 2
|
Cultivar |
Premature
heading
(%) |
Occurrence of
wilting |
Leaf
senescence
at maturing |
Cold tolerancez
|
Low tem. germi-
nationy
(%) |
Viviparous germi-
nationx
(%) |
Seedling
stage
(1-9) |
Heading
delay
(day) |
Grain
fertility
(%) |
Phenotypic
acceptability
(1-9) |
|
Drimi9ho |
0.0±0.0ns w
|
Strongns
|
Latens
|
4±0.3ns
|
8±1.3**
|
35.3±2.5**
|
6±0.8ns
|
79.5±1.8**
|
42.4±3.2**
|
|
Cheongcheong |
0.0±0.0 |
Strong |
Late |
4±0.8 |
14±1.5 |
21.8±1.8 |
5±1.2 |
80.1±1.6 |
51.4±3.8 |
Table 3Lodging-related traits of ‘Drimi9ho’ and performance under field conditions.
Table 3
|
Cultivar |
Plant heightz
(cm) |
Third internode length
(cm) |
Breaking strength
(g) |
Lodging
index |
|
Drimi9ho |
74.7±5.3** y
|
08.3±1.2**
|
1,142.0±3.2**
|
163±5.3**
|
|
Cheongcheong |
92.8±3.8 |
10.2±0.8 |
0994.7±3.4 |
212±4.8 |
Table 4Evaluation of ‘Drimi9ho’ resistance to blast disease across different regions.
Table 4
|
Cultivar |
Leaf blast disease (0-9)z
|
|
Neck blast disease |
|
|
|
No. of tested sites (14) |
Percentage of diseased panicle (%) |
|
|
|
Ry
(0-3) |
M
(4-6) |
S
(7-9) |
M
ean |
Daegu |
Gunwi |
Chilgok |
M
ean |
|
Drimi9ho |
2 |
1 |
0 |
3.6±1.1ns x
|
|
2.3±1.3 |
1.8±1.3 |
2.0±1.0 |
2.0±0.2ns
|
|
Cheongcheong |
2 |
1 |
0 |
3.0±1.2 |
|
1.5±1.1 |
2.1±1.2 |
1.8±1.1 |
1.8±0.3 |
Table 5Resistance evaluation of ‘Drimi9ho’ against bacterial blight, viral diseases, and insect pests.
Table 5
|
Cultivar |
Bacterial blight |
|
Virus disease |
|
Insect pests |
|
|
|
|
K1
|
K2
|
K3
|
K3a
|
RSVz (%) |
RDV (%) |
RBSDV (%) |
BPH |
SBPH |
WBPH |
|
Drimi9ho |
Ry
|
S |
S |
S |
|
R (15.8±2.3)x
|
R (18.6±2.8) |
R (45.5±2.4) |
|
R |
R |
R |
|
Cheongcheong |
R |
S |
S |
S |
|
R (16.3±1.8) |
R (22.4±1.5) |
S (81.4±2.8) |
|
R |
R |
R |
Table 6Evaluation of physicochemical traits affecting grain appearance and quality in ‘Drimi9ho’.
Table 6
|
Cultivar |
Brown rice |
Trans-
lucency
(1-9) |
White
core
(0-9) |
White
belly
(0-9) |
Alkali
digestive
value
(1-7) |
Protein
content
(%) |
Amylose
content
(%) |
Palatability of
cooked rice
(-3-+3) |
Length
(mm) |
Width
(mm) |
Thickness
(mm) |
|
Drimi9ho |
5.12±0.8nsz
|
2.62±1.2ns
|
1.82±1.3ns
|
1±0.0**
|
0±0.0**
|
0±0.0**
|
6.2±0.2ns
|
5.1±1.3**
|
18.6±1.5**
|
0.21±0.1**
|
|
Cheongcheong |
5.08±1.1 |
2.65±1.1 |
1.76±1.1 |
2±0.0 |
1±0.0 |
1±0.0 |
6.7±0.2 |
6.9±1.2 |
17.8±1.3 |
0.12±0.3 |
Table 7Analysis of milling characteristics and associated factors in ‘Drimi9ho’.
Table 7
|
Cultivar |
Milling recovery ratio (%) |
Head rice milling
recovery ratio (%) |
|
Brown/rough |
Milled/brown |
Milled/rough |
Head rice |
|
Drimi9ho |
82.5±2.5** z
|
89.3±3.2**
|
75.3±1.2**
|
91.6±2.5**
|
68.4±3.4**
|
|
Cheongcheong |
81.3±3.4 |
88.4±2.5 |
72.5±3.4 |
89.8±2.3 |
64.2±3.5 |
Table 8Evaluation of ‘Drimi9ho’ yield under diverse environmental conditions by region.
Table 8
|
Culture season |
Region |
Milled rice (kg/10a) |
|
Index |
|
|
|
Drimi9ho (A) |
Cheongcheong (B) |
A/B |
|
Ordinary planting |
Daegu |
562.7±2.8ns z
|
532.8±5.7 |
|
105.6±4.2 |
|
Gunwi |
535.8±3.7**
|
517.6±4.8 |
|
103.5±3.1 |
|
Chilgok |
541.2±5.1**
|
510.8±3.6 |
|
105.9±3.3 |
|
Mean |
546.6±3.8**
|
522.2±5.8 |
|
104.7±4.8 |
References
- 1. Adiredjo AL, Kiswanto I. 2025. Genetic control of resistance mechanisms toward brown planthopper in rice. Plant Breed Biotechnol 13: 1-4.
- 2. Ali S, Ullah MI, Sajjad A, Shakeel Q, Hussain A. 2020. Environmental and health effects of pesticide residues. In: Sustainable agriculture reviews 48: Pesticide occurrence, analysis and remediation vol. 2 analysis. Springer, pp. 311-336.
- 3. Alonso P, Gladieux P, Moubset O, Shih P-J, Mournet P, Frouin J, Blondin L, Ferdinand R, Fernandez E, Julian C. 2019. Emergence of southern rice black-streaked dwarf virus in the centuries-old Chinese yuanyang agrosystem of rice landraces. Viruses 11: 985
- 4. Bandumula N. 2018. Rice production in Asia: Key to global food security. Proc Natl Acad Sci India Sect B Biol Sci 88: 1323-1328.
- 5. Chemists AoOA, Horwitz W. 1960. Official methods of analysis. Association of Official Agricultural Chemists, Washington, DC.
- 6. Fukagawa NK, Ziska LH. 2019. Rice: Importance for global nutrition. J Nutr Sci Vitaminol 65: S2-S3.
- 7. Hoang AT, Zhang H-M, Yang J, Chen J-P, Hébrard E, Zhou G-H, Vinh VN, Cheng J-A. 2011. Identification, characterization, and distribution of southern rice black-streaked dwarf virus in Vietnam. Plant Dis 95: 1063-1069.
- 8. Horgan FG, Garcia CPF, Haverkort F, de Jong PW, Ferrater JB. 2020. Changes in insecticide resistance and host range performance of planthoppers artificially selected to feed on resistant rice. Crop Prot 127: 104963.
- 9. Jeong O-Y, Park H-S, Baek M-K, Kim W-J, Lee G-M, Lee C-M, Bombay M, Ancheta MB, Lee J-H. 2021. Review of rice in Korea: Current status, future prospects, and comparisons with rice in other countries. J Crop Sci Biotechnol 24: 1-11.
- 10. Kim K-M, Park Y-H. 2018. Studies of the life cycle and rearing methods of whitebacked planthopper (Sogatella furcifera Horvath). J Life Sci 28: 357-360.
- 11. Kim M-S, Ouk S, Jung K-H, Song Y, Van Trang L, Yang J-Y, Cho Y-G. 2019. Breeding hybrid rice with genes resistant to diseases and insects using marker-assisted selection and evaluation of biological assay. Plant Breed Biotechnol 7: 272-286.
- 12. Kwak D-Y, Yeo U-S, Lee J-H, Oh B-G, Shin M-S, Ku Y-C. 2007. Mass screening method for rice virus resistance using screen house. Korean J Crop Sci 52: 129-133.
- 13. Park H-S, Baek M-K, Kim W-J, Lee C-M, Ji H, Suh J-P, Jeong O-Y, Cho Y-C, Lee J-H. 2020. Multiple disease resistant early maturing rice cultivar 'IS592BB' with the genetic background of 'Unkwang'. Korean J Breed Sci 52: 473-484.
- 14. Park H-S, Kim K-Y, Kim W-J, Jeung J-U, Lee J-H, Noh T-H, Baek M-K, Shin W-C, Kim J-J, Jeong J-M. 2016. Development of japonica rice lines with four bacterial blight resistance genes using phenotypic and marker-assisted selection. Korean J Breed Sci 48: 140-158.
- 15. Reddy BS, Chaudhari B, Sowmya C, Dawane P. 2022. Evaluation of insecticides for the management of brown plant hopper and white backed plant hopper in rice crop. Biol Forum Int J 14: 511-517.
- 16. Rice INfGEo.1996. Standard evaluation system for rice. RRI, International Rice Research Institute.
- 17. Rural Development Administration (RDA).2017. Manual for rice standard cultivation. RDA, Jeonju, Korea.
- 18. Rural Development Administration (RDA).2018. Report of new cultivars development and research in summer crop. pp. 3-162.
- 19. Seo J, Lee S-M, Han J-H, Shin N-H, Koh H-J, Chin JH. 2019. Identification of yield and yield-related quantitative trait loci for the field high temperature condition in backcross populations of rice (Oryza sativa L.). Plant Breed Biotechnol 7: 415-426.
- 20. Shin E, Kang Y-G, Jeong S-Y, Na J-H, Koo H-N, Kim G-H. 2021. Monitoring of resistant-associated point mutations in the migratory planthoppers Nilaparvata lugens and Sogatella furcifera (Hemiptera: Delphacidae) in Korea, 2020. Korean J Pestic Sci 25: 324-332.
- 21. Yangon M. 2011. Population fluctuations of brown plant hopper nilaparvata lugens stal. and white backed plant hopper sogatella furcifera horvath on. J Entomol 8: 183-190.
- 22. Yuan H. 2012. Effects of temperature on functional response of Anagrus nilaparvatae Pang et Wang (Hymenoptera: Mymaridae) on the eggs of whitebacked planthopper, Sogatella furcifera Horváth and brown planthopper, Nilaparvata lugens Stål. J Integr Agric 11: 1313-1320.
- 23. Zhou G, Xu D, Xu D, Zhang M. 2013. Southern rice black-streaked dwarf virus: A white-backed planthopper-transmitted fijivirus threatening rice production in Asia. Front Microbiol 4: 270