적요
Male sterility is used to mass-produce F1 hybrid seeds in pepper (Capsicum annuum L.). In particular, the development of genetic male sterility (GMS)-linked molecular markers may play a crucial role in hybrid breeding of pepper. To date, approximately 20 GMS genes have been identified in pepper. Among these, several molecular markers for the ms3 gene have been developed in previous studies; however, they are not completely linked and thus have limitations for use in selection. Therefore, in the present study, we aimed to develop molecular markers for ms3 selection using single-nucleotide polymorphism (SNP)-based high-resolution melting (HRM) analysis. Chi-square test was conducted using three F2 segregating populations, and the results confirmed a 3:1 segregation ratio between male-fertile and male-sterile plants. A total of 128 primer sets were designed by selecting SNPs near the ms3 gene, and 25 HRM markers were successfully developed. Using 420 individuals from the F2 segregating population ‘GMS3,’ a high-density genetic linkage map of pepper chromosome 1 was constructed, with eight HRM markers found to be co-segregated with the ms3 gene. Subsequent experiments using various plant materials validated these eight markers, ultimately identifying two HRM markers, HRM119655681 and HRM135273656, for the final selection. These two markers showed co-segregation between the phenotype and genotype of ms3 across all plant materials used in the study. The markers developed in this study are expected to be effective for maternal line development and large-scale F1 hybrid seed production using ms3 in pepper.
서언
고추 웅성불임성은 크게 두 가지로 나뉜다(
Shifriss 1997). 하나는 핵 내 유전자에 의해 결정되는 유전자적 웅성불임성(genetic male sterility, GMS)이며, 다른 하나는 미토콘드리아에 존재하는 웅성불임 유전자에 의해 발생하는 세포질 웅성불임성(cytoplasmic male sterility)이다(
Akter et al. 2016,
Dhaliwal & Jindal 2014). 최근 기후 변화로 고추 품종의 수명이 짧아지면서, 상대적으로 빠르게 모계를 만들 수 있는 GMS를 사용하는 경우가 많아지고 있다. 채종포에서 GMS의 모계는 표현형 분리가 1:1로 가임과 불임으로 분리되며, 꽃의 표현형을 통해 가임 개체를 제거해야 한다(
Rao et al. 1990). 꽃이 피기 전에 불임 개체와 가임 개체를 구분할 수 있는 GMS 유전자 연관 분자표지가 개발된다면 노동력을 크게 절감할 수 있을 것이다(
Jindal et al. 2019).
현재까지
ms1~ms
15, msc-1~msc-3,
msk를 포함하여 최소 20개 이상의 고추 유전자적 웅성불임성이 보고되었다(
Wang & Bosland 2006). 그중
msk는 국내 건고추 품종 개발에,
ms1은 파프리카 품종 개발에,
ms3는 유럽 및 국내 고추 품종 개발에 사용되고 있다. 특히 일부 유전자에 대해서는 후보유전자가 탐색되었다. 먼저
ms1 유전자는 고추 5번 염색체에 위치하였으며,
CA05g06780 유전자가 유력한 후보유전자였는데, 애기장대(
Arabidopsis thaliana)의
MALE STERILITY1 (
MS1) 유전자와 상동유전자였다(
Jeong et al. 2018,
Wilson et al. 2001). 이 유전자는 화분과 융단세포의 발달을 조절하는 PHD-type 전사 조절 인자의 기능을 하는 것으로 보고되었다(
Ito & Shinozaki 2002,
Ito et al. 2007). 다음으로
msc-1 유전자는 2번 염색체에 위치하였으며,
Capana02g002096 유전자가 강력한 후보유전자로 보고되었다(
Cheng et al. 2018). 이 후보유전자는 bHLH 전사인자를 암호화하고 있는
AtDYT1 유전자와 상동유전자였고, 초기 융단세포 발달에 관여한다고 보고되었다(
Zhang et al. 2006). 그리고
msc-2 유전자는 5번 염색체 상에 존재했는데,
AtMS1 유전자와 상동인
Capana05g000766가 후보유전자였다(
Cheng et al. 2020). 또한
msc-3 유전자는 10번 염색체 상에 존재하였으며, MYB80 전사인자를 암호화하고 있는
Capana10g000198 유전자에 163-bp 삽입이 일어나 웅성불임성이 야기된다고 보고되었다(
Dong et al. 2023). 마지막으로
msk 유전자도 10번 염색체에 있었는데,
AtMYB80 유전자와 상동인 T459_24638 (
CaMYB80)를 후보유전자로 선발하였다(
Jeong et al. 2025). 이 웅성불임 유전자에도 163-bp의 삽입이 발견되었는데,
msc-3과
msk 유전자는 하나의 공통 조상의 유전자로부터 유래한 것으로 보고하였다(
Jeong et al. 2025). MYB80 전사인자는 다양한 작물에서 융단조직 및 화분 발달의 조절자로서 작용하며 기능적으로 잘 보전되어 있는 것으로 보고되었다(
Phan et al. 2011,
2012).
하지만
ms3 유전자에 대한 연구는 아직 미흡한 실정이다. 고추 유전자적 웅성불임
ms3는 X-ray 처리에 의한 돌연변이 유도로 만들어졌다고 보고되었다(
Daskaloff 1968). 이후
ms3 유전자와 연관된 GMS3-CAPS 분자표지가 개발되었고, 1번 염색체에 위치한 것으로 확인되었다(
Lee et al. 2010b). 또한 다른 연구에서 HPGMS2 (cleaved amplified polymorphic sequence, CAPS) 및 HPGMS3 (dCAPS) 분자표지가 개발되었지만 이들 분자표지는
ms3 유전자좌와 3.83 centiMorgan (cM) 떨어져 있는 것으로 나타났다(
Naresh et al. 2018). 이는 보다 정밀한 선발을 가능하게 할 새로운 분자표지 개발이 필요하며 이를 해결하기 위해 추가적인 연구와 분석이 요구된다.
따라서 본 연구의 목적은 next-generation sequencing (NGS) resequencing 기반으로 탐색된 SNP를 분자표지로 개발하여 이를 활용해 고추 1번 염색체의 ms3 유전자좌에 고밀도 유전자 지도를 작성하고 더 정확하게 선발할 수 있는 SNP 분자표지를 개발하는 것이다.
재료 및 방법
식물재료 및 표현형 검정
2023년도에 분자표지 개발을 위한 F
2 분리집단 420개체를 육성하였다. F
2 분리집단은 가임(
Ms3ms3)과 불임(
ms3ms3)이 1:1로 분리되는 집단에서 이형접합 가임개체(
Ms3ms3)를 선발한 후 자가수정을 통해 채종된 종자였다. 개발된 분자표지의 유효성 검정을 위해 2024년도에 BC
1, F
1, F
2 등을 포함해 총 527개체를 육성하였다(
Table 1).
표현형 분석과 유전자형 분석을 위한 DNA 추출에 사용된 식물재료의 재배는 전북대학교 농업생명과학대학의 부속농장 유리 온실에서 수행되었다. 종자 발아는 온도 28℃로 유지되는 생장실에서 이루어졌으며, 파종 후 육묘 온도는 고온성 작물인 고추 특성을 고려하여 평균 27℃를 유지하였다.
표현형 검정은 작물의 개화 시기와 화분 방출 시기에 맞춰 오전에 수행하였다. 수술에 화분이 육안으로 확인되면 해당 개체를 웅성가임(MF)으로, 화분이 없거나 수술이 보라색인 경우 웅성불임(MS)으로 판단하였다(
Fig. 1). 개체당 2~3개의 꽃을 3회 조사하였고, 화분이 육안으로 확인되지 않을 경우 검은색 종이에 수술을 문질러 화분 존재 여부를 확인하였다.
분자표지 검정을 위해서 각 고추 식물의 잎에서 DNA를 추출하였다. 고추 식물에서 2-3개의 어린 잎을 사용하였고 DNA 추출은
Eun et al. (2016)의 방법을 사용하였다. 추출된 DNA의 농도는 BioDrop Lite (Biochrom, England)를 사용하여 20 ng⋅μL
-1로 조정하여 이후 실험에 사용하였다.
선행 연구에서 개발된
ms3 연관 분자표지의 물리적 위치를 기반으로 주변의 SNP를 선발하였다(
Geum 2019,
Lee et al. 2010b). 선행 연구에서 개발된 분자표지도 본 연구에서 사용하였다(
Supplementary Table 1). 새롭게 개발된 분자표지는 기존의 차세대 염기서열 재분석(NGS resequencing) 데이터에서 탐색된 SNP를 사용하였다(
Jeong et al. 2018). 프라이머 설계는 Primer 3 Input ver.0.4.0을 사용하여 수행되었다. 고유한 서열을 가진 특이적 분자표지를 개발하기 위해 amplicon 염기서열을 NCBI 데이터베이스에 있는 고추 참조유전체[
Capsicum annuum cv. CM334 ver.1.6; GCA_000512255.2 (ASM51225v2)]에 BLAST 분석을 수행하여 특이적인 것만 분자표지로 개발하였다. 총 128개의 high-resolution melting (HRM) 분석용 프라이머를 설계하였다(
Supplementary Table 2). HRM 뒤 숫자는 SNP의 고추 1번 염색체 상 물리적 위치(bp)를 표시한 것이다.
HRM 분석은
Lee et al. (2022)의 방법대로 수행하였다. 유전자형 분석은 LightCycler
® 96 software ver.1.1 (Roshe Holding AG, Basel, Switzerland)를 이용하여 분석하였다. 유전자형은 세 가지 유형으로 분류하였다. 동형접합체인(homozygous) 웅성가임(
Ms3Ms3, F), 이형접합체인(heterozygous) 웅성가임(
Ms3ms3, H), 그리고 웅성불임(
ms3ms3, S)으로 구분하였다(
Supplementary Fig. 1).
유전자지도는 JoinMap
® 4.1 software (Kyazma
® B.V., Netherlands)을 사용하여 작성하였다. 분자표지 간의 유의미한 연관성을 가진 연관군을 식별하기 위해 LOD 임계값을 3.0으로 설정하였다. 유전자 간의 거리는
Kosambi (2017) mapping function을 사용하여 계산하였다. 유전자지도는 MapChart ver. 2.1 software (
Voorrips 2002)를 사용하여 도식화하였다.
결과 및 고찰
표현형 분리
2023년도와 2024년도에 육성된 두 개의 F
2 분리집단(GMS3와 T-selfing)의 표현형 분리비가 멘델의 유전 법칙에 따라 3:1 비율로 분리되는지 검증하기 위해 적합도 검정(Χ
2 test)을 실시하였다(
Table 2). 그 결과, 모든 카이검정값이 0.05% 기준의 임계치인 3.84를 초과하지 않았고, 통계적 유의성을 판단하는
p-value 값이 기준값 0.05보다 컸다. 이는 관찰된 분리비가 통계적으로 유의미한 차이를 보이지 않는 것을 의미한다. 즉 웅성가임과 웅성불임의 분리비가 3:1의 분리비에 일치함을 의미한다. 따라서, 본 연구에서 사용된 고추 유전자적 웅성불임성이 이전 연구 결과와 동일하게 단일 열성유전자(
ms3)에 의해 나타난다는 것을 확인하였다(
Lee et al. 2010b,
Naresh et al. 2018).
우선 기존 선행연구에서 탐색된
ms3 연관 분자표지의 염기서열을 NCBI 데이터베이스에서 고추 참조유전체[
Capsicum annuum cv. CM334 ver.1.6; GCA_000512255.2 (ASM51225v2)]에 BLAST 분석하여 물리적 위치를 탐색하였다. 그 결과,
ms3 유전자의 위치가 고추 1번 염색체의 100 Mbp에서 145 Mbp 사이로 추정되었다. 따라서 이 범위 내에서 NGS resequencing에서 탐색된 SNP 중 128개를 선발하고 HRM 분석용 프라이머를 설계하였다(
Supplementary Table 2). HRM 분석을 위해 여러 프라이머를 시험하여 다형성이 확인되는 분자표지를 선별하였다. 그 결과, 총 128개 중 25개의 HRM 분석(HRM101096843, HRM102893339, HRM102897000, HRM102902682, HRM102911758, HRM103087926, HRM103088013, HRM103255158, HRM103275816, HRM103292478, HRM103451796, HRM103480835, HRM103480863, HRM104212623, HRM104215541, HRM104224853, HRM105020091, HRM117868894, HRM119042778, HRM119655681, HRM119764966, HRM121624913, HRM129287831, HRM135273656, HRM142660665)에서 뚜렷한 다형성이 확인되어 이후 실험에 HRM 분자표지로 사용하였다(
Supplementary Fig. 1). 개발된 HRM 분자표지는 3가지 유형의 분리 유형을 보였으며, 이와 같은 유형들은 F (homozygous male-fertile), H (heterozygous male-fertile), S (male-sterile)로 분류하였다. 25개를 제외한 프라이머 조합에서는 다형성이 없거나, 두 개 이상의 단편이 증폭되어 유전자형을 구별하는데 어려움이 있어 이후 실험에서 제외하였다(
Supplementary Table 2).
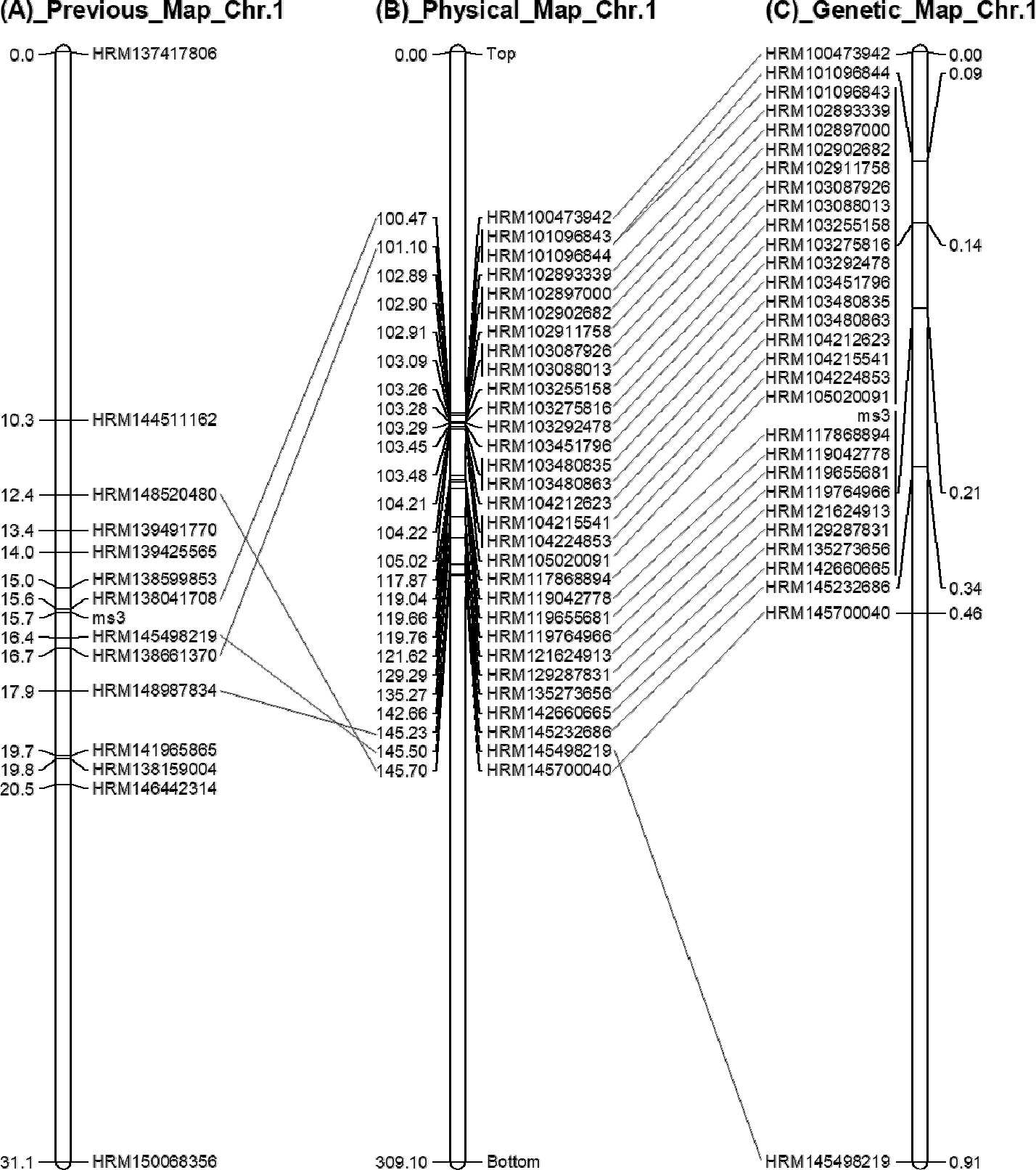
이전 연구에서 14개의 HRM 분자표지를 이용하여 31.1 cM 범위의 유전자지도를 작성하였다(
Fig. 2A,
Geum 2019). 이전 연구에서 사용된 HRM 분자표지는 고추 참조유전체 CM334 ver.1.55 (
Kim et al. 2014)를 기반으로 개발된 것인데, 새롭게 공개된 고추 참조유전체 CM334 ver.1.6을 이용하여 물리적 위치를 다시 분석한 결과 100~145 Mbp에
ms3 유전자가 있을 것으로 분석되었다(
Fig. 2B). 따라서 이 범위 내에 위치하는 128개의 SNP에 대해 프라이머를 설계하였고(
Supplementary Table 2), 그중 25개의 HRM 분자표지를 개발하였다(
Fig. 2B,
Supplementary Fig. 1). 본 연구에서 개발된 25개의 HRM 분자표지와 기존 연구에서 개발된 5개의 HRM 분자표지를 이용하여 F
2 분리집단 420개 개체에 대해 유전자형을 분석하였다. 그 결과를 이용하여 고추 1번 염색체의 고밀도 유전자지도를 구축하였다(
Fig. 2C). 고추 1번 염색체의 물리적지도와 유전자지도를 비교해 본 결과, 분자표지의 순서가 거의 일치하여 ver.1.55에 비해 ver.1.6의 물지적지도가 많이 개선된 것으로 생각된다(
Fig. 2,
Kim et al. 2017).
30개의 HRM 분자표지를 이용하여 F
2 분리집단 420개 개체를 분석한 결과, 표현형과 유전자형의 결과가 상이한 5개의 재조합체(개체번호 52번, 94번, 107번, 118번, 153번)를 찾을 수 있었다(
Supplementary Table 3). 그중 8개의 분자표지(HRM117868894, HRM119042778, HRM119655681, HRM119764966, HRM121624913, HRM129287831, HRM135273656, HRM142660665)에서는 표현형과 유전자형이 완전히 일치하였다(
Supplementary Table 3). 이 결과를 바탕으로
ms3 유전자의 추정 범위를 117 Mbp에서 142 Mbp로 좁힐 수 있었다. 하지만 여전히 해당 범위가 너무 넓어 새로운 식물재료로 이후 추가 실험을 수행하였다.
재조합체를 탐색하기 위해
ms3 유전자와 가장 가깝게 연관된 8개의 분자표지에 대해 추가 실험을 수행하였다. 2024년도에 재배한 BC
1, F
1, F
2 집단(T-selfing1)을 포함한 302개 개체를 대상으로 HRM 분석을 진행하였다(
Table 1). ‘1011’ BC
1 집단에서는 웅성가임과 불임이 8개체와 9개체로 분리되었는데, 웅성가임의 유전자형은 모두 이형접합형(
Ms3ms3, H)이었고, 불임의 유전자형은 모두
ms3ms3 (S)로 나타나 재조합체가 없었다. ‘1S’, ‘T1’, ‘T2’ F
1 집단에서는 예상대로 모두 웅성가임 표현형을 보였고, 분자표지의 유전자형도 모두 이형접합형(H)이었다. 하지만 ‘T3’ F
1 집단에서 재조합형을 보이는 분자표지(HRM119764966) 한 개를 찾았다(
Table 3). 그리고 ‘T-selfing1’ F
2 분리집단에서 7개의 재조합 식물체를 찾았고, 재조합형을 보이는 2개의 분자표지(HRM121624913와 HRM129287831)를 추가로 탐색하였다(
Table 3). 분석 결과를 바탕으로 재조합형을 보이는 3개의 분자표지를 제외하고, 나머지 5개의 분자표지(HRM117868894, HRM119042778, HRM119655681, HRM135273656, HRM142660665)를 이후 실험에 계속 사용하였다. 이 결과를 바탕으로
ms3 유전자의 추정위치는 117~119 Mbp 또는 129~142 Mbp 범위로 좁혀졌다(
Table 3).
선발된 5개의 HRM 분자표지의 적용성 검정을 위해 ㈜하나종묘에서 제공받은
ms3 유전자형을 알고 있는 50개 개체를 분석하였다(
Supplementary Table 4). 해당 개체는 실제 육종 프로그램에서 활용 중인
ms3 계통의 일부 재료로 개발된 분자표지의 적용성을 검증하기 위해 사용되었다. 분석 결과, HRM142660665에서는 다수의 개체에서 실제 유전자형과 분자표지의 유전자형이 일치하지 않아서 해당 분자표지가 웅성불임 선발 분자표지로 활용되기에 부적합하다고 판단하였다. 또한, HRM117868894, HRM119042778에서도 특정 계통에서 실제 유전자형과 분자표지의 유전자형이 일치하지 않는 것을 확인하여 이들 분자표지 또한 부적합하다고 판단하였다. 반면, HRM119655681과 HRM135273656의 경우, 50개 개체 모두 실제 유전자형과 분자표지의 유전자형이 완전히 일치하여 해당 2개의 분자표지는 선발에 효과적일 것으로 생각되었다.
최종 선발된 2개의 분자표지(HRM119655681과 HRM135273656)의 선발 효율을 검정하기 위하여 ‘T-selfing2’ F
2 분리집단 225개 개체를 이용하여 분석하였다(
Table 1). 분석 결과, 모든 개체에서 표현형과 유전자형이 완전히 일치하였다(
Table 4). 이러한 결과는 두 분자표지가
ms3 유전자와 매우 가깝게 연관되어 있다는 것을 뜻하며 선발 분자표지로서의 높은 정확성을 가지고 있다고 생각된다.
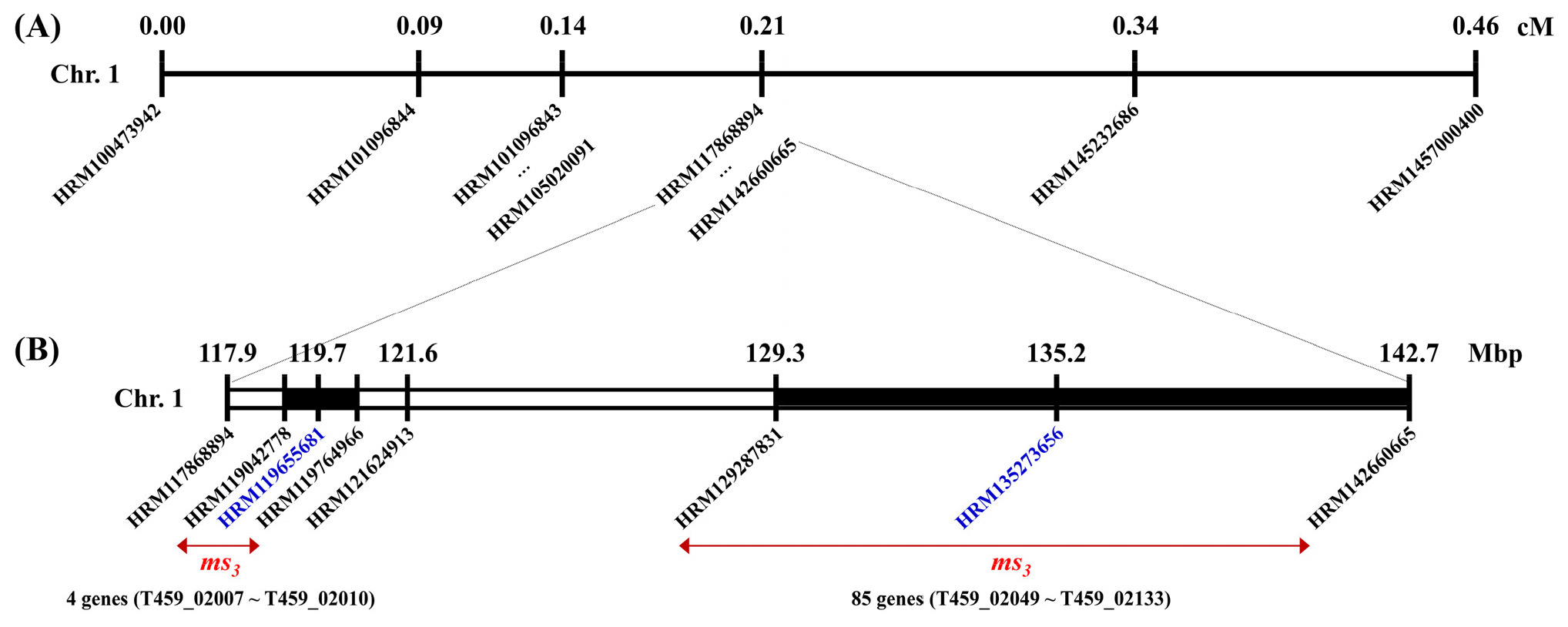
지금까지의 결과를 정리해 보면, 재조합체가 하나도 나타나지 않은 분자표지는 2개로 HRM119655681과 HRM135273656이었다(
Tables 3 and
4,
Supplementary Tables 3 and 4). 이를 기준으로
ms3 유전자는 고추 1번 염색체의 119.0~119.8 Mbp 또는 129.3~142.7 Mbp 범위에 존재할 것으로 추정된다(
Fig. 3). 먼저 119.0~119.8 Mbp 범위에는 4개의 유전자(T459_02007~T459_02010)가 있었는데, 엽록체 리보솜 단백질 유전자 3개와 개화 조절 단백질 유전자 1개였는데, 이들은 웅성불임성과 크게 관련이 없는 유전자였다(
Supplementary Table 5). 이들 유전자들의 코딩 서열을 Sanger sequencing을 통해 분석한 결과, 웅성가임과 웅성불임 개체의 염기서열에 유전적 변이가 존재하지 않았다. 또한, 129.3~142.7 Mbp 범위에는 85개의 유전자(T459_02049~T459_02133)가 밀집되어 있어 정확한 후보 유전자 선정을 위해서는 후속 분석이 요구된다. 진핵생물의 염색체에서 재조합률은 말단소체(telomere) 부근이 동원체(centromere) 부근보다 더 높은 것으로 알려져 있다(
Nachman 2002). 따라서 말단소체에서는 재조합률이 상대적으로 높기 때문에 물리적지도의 범위가 좁은 반면, 동원체 부근에서는 재조합률이 상대적으로 낮기 때문에 물리적지도의 범위가 더 넓게 나타난다. 본 연구에서
ms3 유전자의 물리적 위치가 말단소체 부근이 아니라 동원체 부근이기 때문에 물리적 거리가 넓게 분포함에도 재조합 개체가 상대적으로 적게 나타난 것으로 생각된다.
ms3 유전자의 범위를 더 좁히고 후보유전자의 수를 줄이기 위해서는 개체수를 많이 늘려 추가 실험을 수행해야 할 것으로 생각된다.
적요
우리나라에서 재배되는 모든 고추(Capsicum annuum L.) 품종은 잡종강세를 이용한 일대잡종(F1) 종자이다. 고추에서는 일대잡종 종자를 대량 생산하기 위해 웅성불임성(male sterility)을 이용하고 있다. 특히, 유전자적 웅성불임성(genetic male sterility, GMS) 연관 분자표지 개발은 고추 일대잡종 종자 대량생산의 효율성을 향상시키는 데 중요한 역할을 할 수 있다. 현재까지 고추에서 GMS 관련 유전자는 ms1~ms15, msc-1~msc-3, msk를 포함하여 최소 20개 이상이 알려져 있다. 그중 ms3 유전자의 경우, 이전 연구에서 몇몇 분자표지가 보고되고 있지만 완전 연관 분자표지가 아니기 때문에 선발용으로 사용되기에는 한계가 있다. 따라서 본 연구에서는 고추 참조유전체를 활용하여 단염기다형성(single nucleotide polymorphism, SNP)을 기반으로 한 HRM (high-resolution melting) 분자표지를 이용함으로써 ms3 선발용 분자표지를 개발하고자 한다. 분자표지 개발과 적용성 검정을 위해 여러 식물재료를 육성하였고, F2 분리집단에 대해서는 적합도 검정을 수행하였는데 그 결과 웅성가임과 웅성불임의 분리비가 3:1임을 확인하였다. 이전 연구를 기반으로 ms3 유전자 주변의 SNP를 기반으로 총 128개의 프라이머를 설계하였으며 그중 25개의 HRM 분자표지를 개발하였다. F2 분리집단 420개 개체를 이용하여 고추 1번 염색체의 고밀도 유전자지도를 작성하였고, 그 결과 8개의 분자표지에서 ms3 유전자와 재조합체가 나타나지 않았다. 그 이후 여러 식물재료를 활용하여 8개 분자표지에 대해 추가실험 한 결과, 최종적으로 2개의 HRM 분자표지(HRM119655681과 HRM135273656)를 선발할 수 있었고, 이들 분자표지는 본 실험에서 사용된 모든 식물재료에 대해 표현형과 유전자형이 완전히 일치하였다. 본 연구에서 개발된 분자표지는 고추에서 ms3를 이용한 모계육성 및 F1 종자 대량생산에 효율적으로 사용될 수 있을 것으로 생각된다.
보충자료
본문의 Supplementary Tables 1-5와 Supplementary Fig. 1은 한국육종학회지 홈페이지에서 확인할 수 있습니다.
사사
본 연구는 농촌진흥청 차세대바이오그린21 식물분자육종사업(RS-2018-RD008279)의 지원으로 수행되었습니다.
Fig. 1Phenotypes of male-fertile (A) and male-sterile (B) flower in chili pepper. (A) Homozygous (Ms3Ms3) and heterozygous (Ms3ms3) male-fertile; (B) Male-sterile (ms3ms3).

Fig. 2Comparison of chromosome 1 among the previous genetic map (A,
Geum 2019), a physical map based on CM334 ver. 1.6 (B) and a genetic linkage map with 30 HRM markers (C). Bar left number in (A)_Previous_Map_Chr.1, genetic map position (cM); Bar left number in (B)_Physical_Map_Chr.1, physical map position (Mbp); Bar right number in (C)_Genetic_Map_Chr.1, genetic map position (cM); Bar loci name, SNP marker name; Same loci were connected with lines.
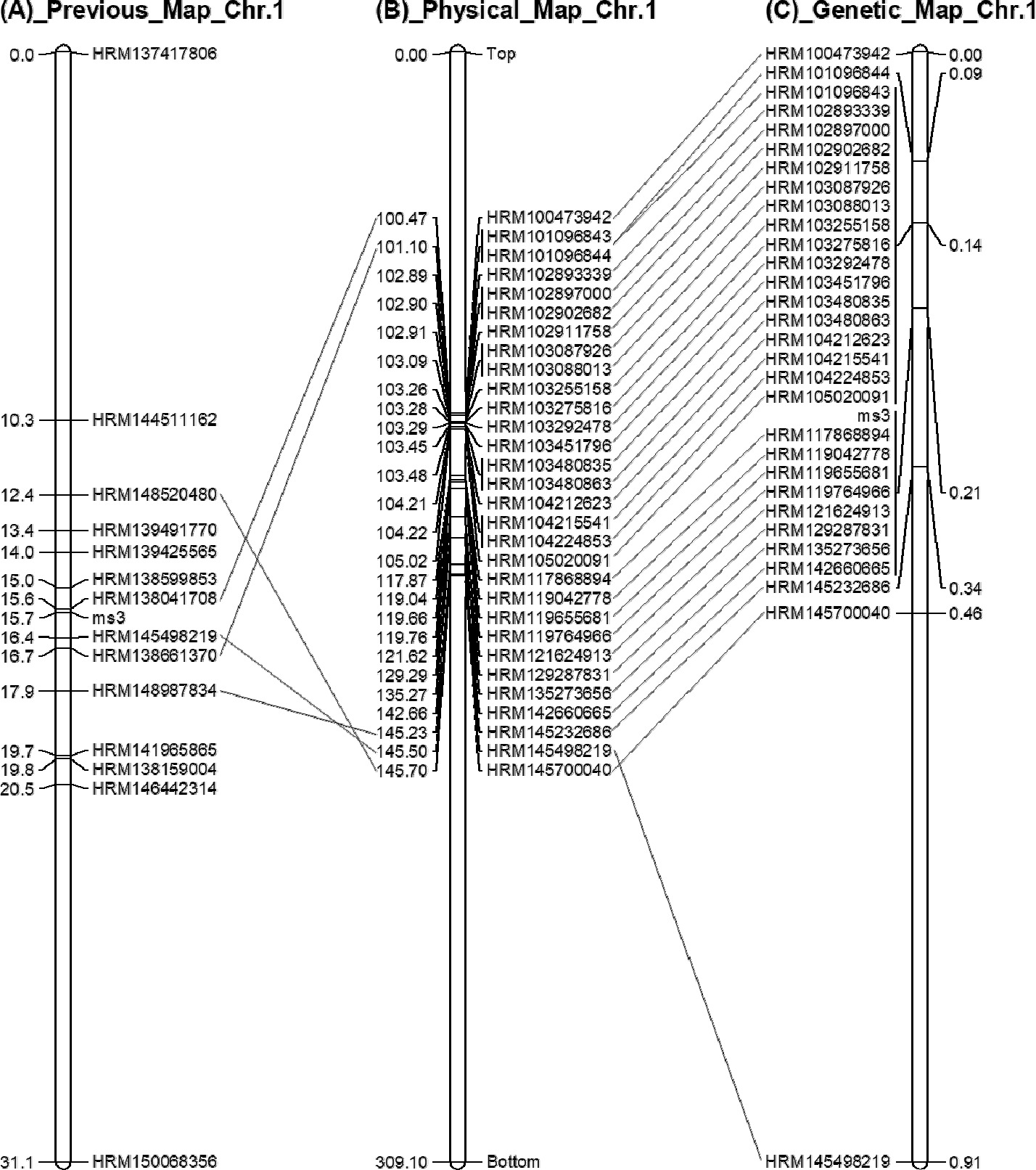
Fig. 3Fine mapping of the ms3 locus in C. annuum. (A) A genetic linkage map around the ms3 locus on chromosome 1 of C. annuum. (B) A physical map of the ms3 locus. Marker name indicates physical position (bp) on chromosome 1. Linkage map distance and physical position are shown upper the marker. Blue-colored markers have no recombinants in this study. Red arrows indicate the possible positions of the ms3 gene.
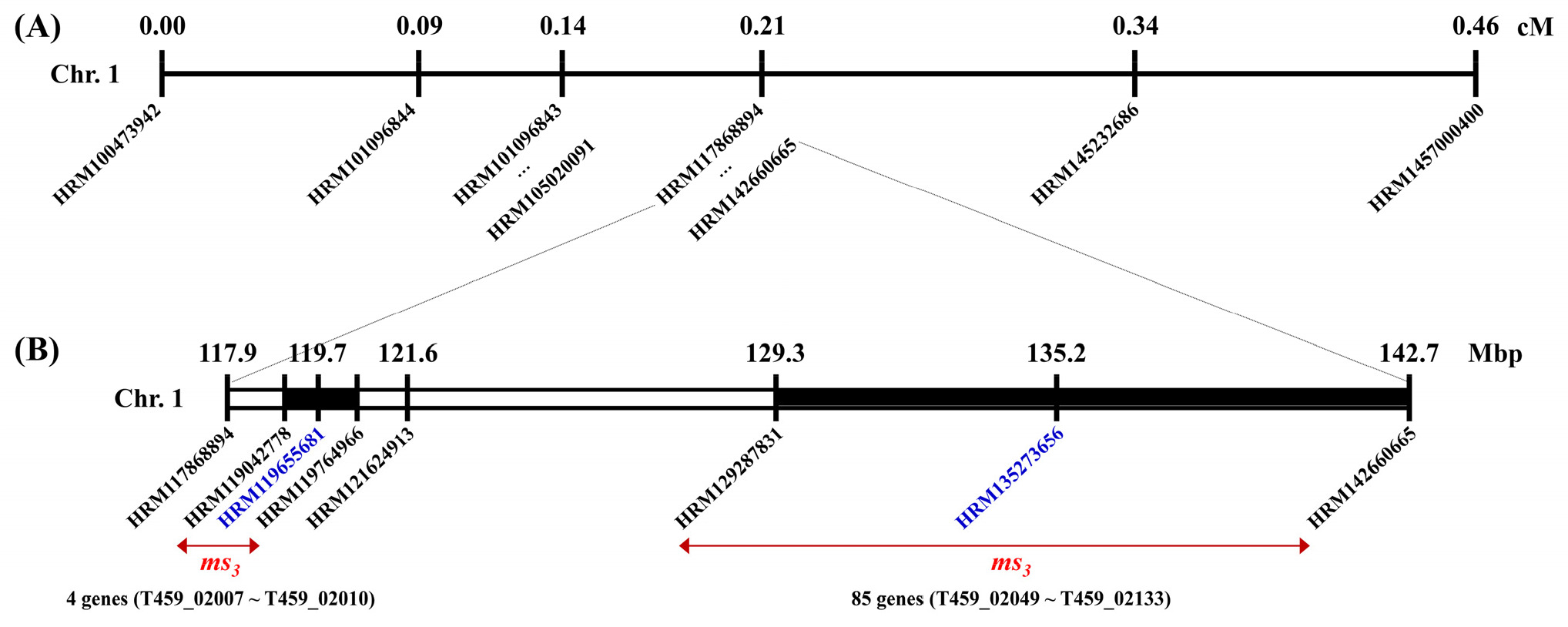
Table 1Plant materials for development and validation of markers linked to ms3 gene.
Table 1
|
No. |
Cultivation year |
Population name |
Generation |
Number of
plants |
Genotype |
Usage |
|
1 |
2023 |
GMS3 |
F2
|
420 |
Segregated (Ms3Ms3, Ms3ms3, ms3ms3) |
Marker development |
|
2 |
2024 |
1S |
F1
|
12 |
Ms3ms3
|
Marker validation |
|
3 |
2024 |
1011 |
BC1
|
17 |
Segregated (Ms3ms3, ms3ms3) |
Marker validation |
|
4 |
2024 |
T1 |
F1
|
5 |
Ms3ms3
|
Marker validation |
|
5 |
2024 |
T2 |
F1
|
5 |
Ms3ms3
|
Marker validation |
|
6 |
2024 |
T3 |
F1
|
5 |
Ms3ms3
|
Marker validation |
|
7 |
2024 |
T-selfing1 |
F2
|
258 |
Segregated (Ms3Ms3, Ms3ms3, ms3ms3) |
Marker validation |
|
8 |
2024 |
T-selfing2 |
F2
|
225 |
Segregated (Ms3Ms3, Ms3ms3, ms3ms3) |
Marker validation |
Table 2Chi-square test for segregation of male-fertile and sterile plants in F2 populations.
Table 2
|
Population |
Generation |
Number of plants |
Expected ratio |
χ2 value |
p-value |
|
|
Total |
Male-fertile |
Male-sterile |
|
GMS3 |
F2
|
420 |
332 |
88 |
3:1 |
3.67 |
0.055 |
|
T-selfing1 |
F2
|
258 |
190 |
68 |
3:1 |
0.25 |
0.615 |
|
T-selfing2 |
F2
|
225 |
165 |
60 |
3:1 |
0.33 |
0.564 |
Table 3Male-fertility phenotype and genotypes of 8 polymorphic HRM markers in recombinant pepper plants.
Table 3
|
Gen.z
|
Plant No. |
Phe.y
|
Genotype of HRM markers |
|
HRM11
7868894 |
HRM11
9042778 |
HRM11
9655681 |
HRM11
9764966 |
HRM12
1624913 |
HRM12
9287831 |
HRM13
5273656 |
HRM14
2660665 |
|
F1
|
T-3-1 |
MF |
Hx
|
H |
H |
Sw
|
H |
H |
H |
H |
|
F1
|
T-3-2 |
MF |
H |
H |
H |
S |
H |
H |
H |
H |
|
F1
|
T-3-3 |
MF |
H |
H |
H |
S |
H |
H |
H |
H |
|
F1
|
T-3-4 |
MF |
H |
H |
H |
S |
H |
H |
H |
H |
|
F1
|
T-3-5 |
MF |
H |
H |
H |
S |
H |
H |
H |
H |
|
|
F2
|
T-selfing1-49 |
MF |
H |
H |
H |
H |
H |
S |
H |
H |
|
F2
|
T-selfing1-141 |
MF |
F |
F |
F |
F |
F |
H |
F |
F |
|
F2
|
T-selfing1-237 |
MF |
H |
H |
H |
H |
H |
S |
H |
H |
|
F2
|
T-selfing1-242 |
MF |
F |
F |
F |
F |
H |
F |
F |
F |
|
F2
|
T-selfing1-137 |
MS |
S |
S |
S |
S |
S |
H |
S |
S |
|
F2
|
T-selfing1-150 |
MS |
S |
S |
S |
S |
S |
H |
S |
S |
|
F2
|
T-selfing1-251 |
MS |
S |
S |
S |
S |
H |
S |
S |
S |
Table 4Validity test of two HRM markers in 225 plants of an F2 population.
Table 4
|
Population |
Generation |
Phenotypez
|
Number of plants |
|
|
HRM119655681y
|
|
HRM135273656 |
|
|
|
F |
H |
S |
F |
H |
S |
|
T-selfing2 |
F2
|
MF |
57 |
108 |
0 |
|
57 |
108 |
0 |
|
MS |
0 |
0 |
60 |
|
0 |
0 |
60 |
References
- 1. Akter S, Huq MA, Jung Y-J, Cho Y-G, Kang K-K. 2016. Application of single nucleotide polymorphism markers for selection of male sterility in crop plants. Plant Breed Biotech 4: 379-386.
- 2. Barka GD, Lee J. 2020. Molecular marker development and gene cloning for diverse disease resistance in pepper (Capsicum annuum L.): Current status and prospects. Plant Breed Biotech 8: 89-113.
- 3. Cheng Q, Li T, Ai Y, Lu Q, Wang Y, Wu L, Liu J, Sun L, Shen H. 2020. Phenotypic, genetic, and molecular function of msc-2, a genic male sterile mutant in pepper (Capsicum annuum L.). Theor Appl Genet 133: 843-855.
- 4. Cheng Q, Wang P, Liu J, Wu L, Zhang Z, Li T, Gao W, Yang W, Sun L, Shen H. 2018. Identification of candidate genes underlying genic male-sterile msc-1 locus via genome resequencing in Capsicum annuum L. Theor Appl Genet 131: 1861-1872.
- 5. Daskaloff S. 1968. A male sterile pepper (C. annuum L.) mutant. Theor Appl Genet 38: 370-372.
- 6. Dhaliwal MS, Jindal SK. 2014. Induction and exploitation of nuclear and cytoplasmic male sterility in pepper (Capsicum spp.): A review. J Hort Sci Biotechnol 89: 471-479.
- 7. Dong J, Hu F, Guan W, Yuan F, Lai Z, Zhong J, Liu J, Wu Z, Cheng J, Hu KA. 2023. 163-bp insertion in the Capana10g000198 encoding a MYB transcription factor causes male sterility in pepper (Capsicum annuum L.). Plant J 113: 521-535.
- 8. Eun MH, Han J-H, Yoon JB, Lee J. 2016. QTL mapping of resistance to the cucumber mosaic virus P1 strain in pepper using a genotyping-by-sequencing analysis. Hortic Environ Biotechnol 57: 589-597.
- 9. Geum B. 2019. SNP marker development and fine mapping for genic male-sterile gene ms3 in chili pepper (Capsicum annuum L.). Master's Thesis Jeonbuk National University. Jeonju. Korea.
- 10. Herath HMSN, Rafii MY, Ismail SI, Nakasha JJ, Ramlee SI. 2021. Improvement of important economic traits in chilli through heterosis breeding: A review. J Hort Sci Biotechnol 96: 14-23.
- 11. Ito T, Nagata N, Yoshiba Y, Ohme-Takagi M, Ma H, Shinozaki K. 2007. Arabidopsis MALE STERILITY1 encodes a PHD-type transcription factor and regulates pollen and tapetum development. Plant Cell 19: 3549-3562.
- 12. Ito T, Shinozaki K. 2002. The MALE STERILITY1 gene of Arabidopsis, encoding a nuclear protein with a PHD-finger motif, is expressed in tapetal cells and is required for pollen maturation. Plant Cell Physiol 43: 1285-1292.
- 13. Jarret RL, Barboza GE, Batista FRC, Berke T, Chou Y-Y, Hulse-Kemp A, Ochoa-Alejo N, Tripodi P, Veres A, Garcia CC, Csillery G, Huang Y-K, Kiss E, Kovacs Z, Kondrak M, Arce-Rodriguez ML, Scaldaferro MA, Szonk A. 2019. Capsicum-An abbreviated compendium. J Amer Soc Hort Sci 144: 3-22.
- 14. Jeong H-J, Jo YD, Park S-W, Kang B-C. 2010. Identification of Capsicum species using SNP markers based on high resolution melting analysis. Genome 53: 1029-1040.
- 15. Jeong K, Choi D, Lee J. 2018. Fine mapping of the genic male-sterile ms1 gene in Capsicum annuum L. Theor Appl Genet 131: 183-191.
- 16. Jeong K, Kim D, Ro N, Lee J. 2025. Development of a molecular marker based on the candidate gene CaMYB80 for genic male sterility msk in pepper (Capsicum annuum L.). Hortic Environ Biotechnol 66: 879-888.
- 17. Jindal SK, Dhaliwal MS, Meena OP. 2019. Molecular advancements in male sterility systems of Capsicum: A review. Plant Breeding 139: 42-64.
- 18. Kim S, Park J, Yeom S-I, Kim Y-M, Seo E, Kim K-T, Kim M-S, Lee JM, Cheong K, Shin H-S, Kim S-B, Han K, Lee J, Park M, Lee H-A, Lee H-Y, Lee Y, Oh S, Lee JH, Choi E, Choi E, Lee SE, Jeon J, Kim H, Choi G, Song H, Lee JK, Lee S-C, Kwon J-K, Lee H-Y, Koo N, Hong Y, Kim RW, Kang W-H, Huh JH, Kang B-C, Yang T-J, Lee Y-H, Bennetzen JL, Choi D. 2017. New reference genome sequences of hot pepper reveal the massive evolution of plant disease-resistance genes by retroduplication. Genome Biol 18: 210
- 19. Kim S, Park M, Yeom SI, Kim YM, Lee JM, Lee HA, Seo E, Kim KT, Kim JH, Cheong K, Shin HS, Kim S, Ryu J, Kim SB, Oh SK, Jo SH, Kim YS, Nam J, Kim BY, Choi D. 2014. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat Genet 46: 270-278.
- 20. Kosambi DD. 2017. The estimation of map distances from recombination values. pp. 125-130. In: Ramaswamy R. (Ed) DD. Kosambi. Springer, New Delhi:
- 21. Lee H-R, An HJ, Yang DC, Choi SH, Kim H-J, Rhee HG, Harn CH. 2012. Development of a high resolution melting (HRM) marker linked to genic male sterility in Capsicum annuum L. Plant Breeding 131: 444-448.
- 22. Lee J, Lee W-P, Han J-H, Yoon JB. 2010a. Development of molecular marker linked to a genic male-sterile gene, msk in chili pepper. Kor J Hort Sci Technol 28: 270-274.
- 23. Lee J, Yoon JB, Han J-H, Lee WP, Kim SH, Park HG. 2010b. Three AFLP markers tightly linked to the genic male sterility ms3 gene in chili pepper (Capsicum annuum L.) and conversion to a CAPS marker. Euphytica 173: 55-61.
- 24. Lee S, Chakma N, Joung S, Lee JM, Lee J. 2022. QTL mapping for resistance to bacterial wilt caused by two isolates of Ralstonia solanacearum in chili pepper (Capsicum annuum L). Plants 11: 1551
- 25. Lozada DN, Bosland PW, Barchenger DW, Haghshenas-Jaryani M, Sanogo S, Walker S. 2022. Chile pepper (Capsicum) breeding and improvement in the "multi-omics" era. Front Plant Sci 13: 879182.
- 26. Nachman MW. 2002. Variation in recombination rate across the genome: Evidence and implications. Curr Opin Genet Develop 12: 657-663.
- 27. Naresh P, Lin S-W, Lin C-Y, Wang Y-W, Schafleitner R, Kilian A, Kumar S. 2018. Molecular markers associated to two non-allelic genic male sterility genes in peppers (Capsicum annuum L.). Front Plant Sci 9: 1343
- 28. Park S-W, An S-J, Yang H-B, Kwon J-K, Kang B-C. 2009. Optimization of high resolution melting analysis and discovery of single nucleotide polymorphism in Capsicum. Hort Environ Biotechnol 50: 31-39.
- 29. Phan HA, Iacuone S, Li SF, Parish RW. 2011. The MYB80 transcription factor is required for pollen development and the regulation of tapetal programmed cell death in Arabidopsis thaliana. Plant Cell 23: 2209-2224.
- 30. Phan HA, Li SF, Parish RW. 2012. MYB80, a regulator of tapetal and pollen development, is functionally conserved in crops. Plant Mol Biol 78: 171-183.
- 31. Rao MK, Devi KU, Arundhati A. 1990. Applications of genic male sterility in plant breeding. Plant Breeding 105: 1-25.
- 32. Ro N, Haile M, Hur O, Geum B, Rhee J, Hwang A, Kim B, Lee J, Hahn B-S, Lee J, Kang B-C. 2022. Genome-wide association study of resistance to Phytophthora capsici in the pepper (Capsicum spp.) collection. Front Plant Sci 13: 902464.
- 33. Semagn K, Babu R, Hearne S, Olsen M. 2014. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): Overview of the technology and its application in crop improvement. Mol Breeding 33: 1-14.
- 34. Shifriss C. 1997. Male sterility in pepper (Capsicum annuum L.). Euphytica 93: 83-88.
- 35. Simko I. 2016. High-resolution DNA melting analysis in plant research. Trends Plant Sci 21: 528-537.
- 36. Voorrips RE. 2002. MapChart: Software for the graphical presentation of linkage maps and QTLs. J Hered 93: 77-78.
- 37. Wang D, Bosland PW. 2006. The genes of Capsicum. HortScience 41: 1169-1187.
- 38. Wilson ZA, Morroll SM, Dawson J, Swarup R, Tighe PJ. 2001. The Arabidopsis MALE STERILITY1 (MS1) gene is a transcriptional regulator of male gemetogenesis, with homology to the PHD-finger family of transcription factors. Plant J 28: 27-39.
- 39. Zhang W, Sun Y, Timofejeva L, Chen C, Grossniklaus U, Ma H. 2006. Regulation of Arabidopsis tapetum development and function by DYSFUNCTIONAL TAPETUM1 (DYT1) encoding a putative bHLH transcription factor. Development 133: 3085-3095.