적요
‘Boramchan’ is a high-yielding, mid-late maturing temperate japonica rice cultivar developed in Korea, while ‘Pecos’ is an early-maturing tropical japonica cultivar from the United States with medium-grain shape characteristics. In this study, quantitative trait loci (QTL) analysis was conducted on yield- and grain-related traits using recombinant inbred lines derived from a cross between ‘Boramchan’ and ‘Pecos’ in order to broaden the genetic diversity of these traits in Korean japonica rice varieties. QTLs for heading date were identified on chromosomes 3 and 6, with Hd6 and Hd1 proposed as candidate genes for this trait. The major heading date gene Hd1 exhibited pleiotropic effects, influencing not only the heading date (HD) but also multiple yield components, including culm length, panicle length, number of spikelets per panicle (NS), 1,000-grain weight (TGW), ratio of ripened grain (RRG), brown/rough rice ratio, and grain yield per plant. The minor heading date gene Hd6 showed a masking effect on HD and NS, depending on the allele type of Hd1. Allelic combinations of Hd1 and Hd6 allowed HD regulation for approximately 30 days. As more functional alleles associated with delayed heading accumulated, the values of yield-related traits gradually increased. Three QTLs for grain-related traits–qGL2, GW5, and qGS10–were identified on chromosomes 2, 5, and 10, respectively. GW5 was confirmed as a major gene with significant effects on phenotypic variation, influencing all grain-related traits, including grain length (GL), width (GW), thickness (GT), length to width ratio, and TGW. qGL2 was associated with GL and colocalized with qNS2, a QTL related to NS. qGS10 was identified as a QTL that affected GW, GT, and TGW. Allelic combinations of qGL2, GW5, and qGS10 influenced not only grain-related traits but also variation in yield-related traits. Combinations carrying the gw5- allele from ‘Boramchan’ generally exhibited favorable characteristics in terms of yield potential. Furthermore, the introduction of qGL2P and qGS10P alleles from ‘Pecos’ contributed to the diversification of grain shape and showed positive effects on NS, RRG, and yield. Novel allele combinations, such as qGL2P-gw5--qGS10B and qGL2P-gw5--qGS10P are expected to enhance the genetic diversity of yield- and grain-related traits in Korean japonica rice varieties.
서언
아시아 재배벼(
Oryza sativa L.)는 형태학적 특성, 서식지와 생태계, 지리적 분포, 재배 문화 등에 따라 크게 인디카(
indica)와 자포니카(
japonica)의 두 가지 아종(subspecies)으로 분류되어 왔다(
Glaszmann 1987,
Wang et al. 2023). 이후 유전적 구조와 전장 유전체 분석을 통해 세분화되어 인디카,
aus, 온대 자포니카(
temperate japonica), 열대 자포니카(
tropical japonica), aromatic 등 5개의 생태형으로 분류되고 있다(
Garris et al. 2005). 인디카 벼는 저위도의 열대와 아열대 지역의 기후에 적합하게 적응하여 중국 남부, 베트남, 태국, 인도 등에서 주로 재배되고, 온대 자포니카 벼는 고위도의 온대지역이나 온도가 낮은 고지대에 적응하여 우리나라를 비롯하여 일본, 중국 동북지역 등 인디카에 비해 다소 제한된 지역에서 재배되고 있다(
Kovach et al. 2007,
Vaughan et al. 2008). 온대 자포니카 벼는 낟알의 길이가 짧고 형태가 둥근 단립종의 단원형 입형 특성을 가지는 것이 많으며 아밀로스 함량이 낮고 밥을 지었을 때 찰성이 있고 끈기가 있는 반면, 인디카 벼는 낟알의 길이가 길고 너비가 얇은 장립종의 세장형 입형 특성을 나타내는 것이 대부분이며 밥의 찰기가 상대적으로 떨어지며 푸석푸석한 느낌을 가지는 경우가 많다(
Calingacion et al. 2014,
Juliano 1979). 우리나라 사람들은 예로부터 밥이 차지며 끈기가 있는 것을 좋아하는데 이로 인해 온대 자포니카형 위주의 벼 육종사업이 지속되어 우리나라에서 재배되는 밥쌀용 벼 품종들은 대부분 온대 자포니카 벼이다(
Park et al. 2018). 온대 자포니카 벼는 인디카에 비해 유전적 다양성이 협소한 것으로 알려져 있다(
Wang et al. 2018). 국내 벼 육종사업에서 비슷한 유전조성을 가지는 온대 자포니카 자원들을 지속적으로 이용하다 보니 유전적 다양성이 부족하게 되고 이는 변화되는 기후에 대한 재배안정성 약화로 이어질 수 있어 이에 대한 개선이 필요하다.
미국 재배 벼의 집단 구조 분석 결과, 인디카보다 자포니카 아종이 대부분을 차지하며, 중단립종의 온대 자포니카보다 중립종 및 장립종의 열대 자포니카 생태형이 더 많이 포함된 것으로 보고되었다(
Lu et al. 2005). 열대 자포니카 벼는 온대 자포니카 벼보다 고온 조건에서 적응성이 높으며, 유전적 다양성이 높은 것으로 알려져 있다(
Singh et al. 2022,
Wang et al. 2023). ‘Pecos’, ‘Cypress’ 등 미국 14개 품종을 우리나라 환경조건에서 농업형질 특성을 검토한 결과, ‘보람찬’에 비해 출수가 빠른 준조생-중만생종의 출수 생태형에 간장과 수장이 길고 수수는 적고 수당립수는 많았으며 등숙률과 간이 수량이 낮은 편이었다(
Park et al. 2022). 외형적 특성에서는 14개 미국 품종 모두 잎과 영에 강모가 없는 매끄러운 특성을 나타냈으며 황금색 영, 자주색 부선 및 주두를 가진 품종들이 존재하여 우리나라 품종과 형태학적으로 다른 특성을 나타냈다. 입형 특성에서 미국 품종은 중립종, 장립종, 초장립종의 다양한 길이를 가지고 있었으며, 중원형과 세장형과 같이 우리나라 자포니카 벼 품종이 가지고 있지 않은 입형 특성을 나타내 우리나라 벼 품종의 입형 다양화를 위한 자원으로 활용될 것으로 기대된다 하였다(
Park et al. 2022).
출수기는 벼가 발아하고 영양생장과 생식생장을 거쳐 이삭이 나오기까지의 기간을 나타내며 지역의 환경조건과 작용하여 재배적응성을 결정하고 수량 및 품질 등 주요 농업형질 특성에 영향을 미치기 때문에 벼 육종에 있어서 중요한 목표 형질이다(
Kim et al. 2018). 우리나라에서는 출수생태에 따라 조생종, 중생종, 만생종으로 구분하며, 생육기간은 조생종은 약 140일, 중생종 160일, 만생종 180일 정도이다(
Kim 2010). 우리나라 벼 출수 생태형별 재배면적을 살펴보면 중만생종이 80%를 넘을 정도로 중만생종 품종에 편중된 벼농사가 이루어지고 있다. 중만생종에 편중된 재배는 수확기 노동력의 과중 문제, 집중 출하에 따른 건조⋅저장 등 수확 후 관리에 어려움을 가지오며 급격한 쌀값 변동에 영향을 줄 수 있다(
Nam et al. 2019). 또한 최근 논의 활용도와 곡물자급률을 높이기 위해 가루쌀과 밀, 소득작물 후작 등 이모작 작부체계가 증가하는 추세이나, 생육기간이 긴 중만생종의 경우 이앙기가 후작물과 재배시기가 겹치기 때문에 이모작 재배에 제한적이다. 이러한 문제를 극복하기 위해 조생종, 중생종, 소득작물 후작용 등 출수 특성이 다양화된 품종 개발에 대한 요구가 증가하고 있다.
벼 낟알의 크기와 형태인 입형은 수량과 품질에 중요한 역할을 하는 형질이다. 우리나라 자포니카 벼 품종들은 대부분 중단립종의 단원형의 매우 협소한 입형 특성을 가지고 있다(
Lee et al. 2020). 협소한 유전적 배경으로 인하여 새롭게 개발되는 육성 계통이나 품종들의 입형은 중단립종의 단원형 위주로 고착화되었다. 우리나라 자포니카 벼의 정체되어 있는 수량성 장벽과 기후변화에 따른 품질저하를 극복하기 위한 기반연구와 더불어 새로운 품질에 대한 소비자의 욕구 충족과 쌀 산업 발전을 위한 사회적 변화를 견인하기 위해서는 우리나라 자포니카 벼 품종의 입형 특성을 다양화 해야 한다 하였다(
Park et al. 2017). 미국품종 ‘Calrose’에서 유래된 것으로 확인된
gs3 대립유전자는 ‘신동진’과 같은 중대립 품종에 도입되어 수량과 품질에 있어서 기존의 국내 품종들이 가지지 못했던 특성을 나타내는 등 유전적 다양성 확충에 기여하였다(
Lee et al. 2020,
Park et al. 2024a,
Park et al. 2024b). 또한
gs3의 도입은 메탄 생성을 저감하고 수량성을 향상시킴으로써 질소 비료 사용을 줄이는 등 온실가스 감축에 효과가 있을 것으로 보고되었다(
Kwon et al. 2023). 이처럼 다른 생태형에서 국내 온대 자포니카 벼로 이입된 새로운 대립유전자가 수량과 품질 관련 형질의 변이 확대 뿐만 아니라 기후변화에 대응하기 위한 유용인자로도 활용될 수 있는 등 육종의 가치 증대에 기여할 수 있다.
본 연구는 우리나라 온대 자포니카 벼 품종과 형태학적으로 차이가 나고 유전적 다양성이 다른 것으로 알려진 미국 품종을 교배모본으로 이용하여 재조합 자식 집단을 육성하고 수량과 입형 관련 형질에 대한 QTL 분석을 수행하였다. 이를 통해 수량과 입형 관련 형질이 다양화된 육종소재를 확보하고, 형질 변이에 대한 유전 양상과 관련 유전자들의 작용을 파악하여 육종사업에 반영함으로써 다양한 입형 특성을 가지면서 수량성이 향상된 자포니카 벼 품종 개발에 기여하고자 수행되었다.
재료 및 방법
시험재료 및 재배방법
시험재료로 ‘보람찬/Pecos’ 재조합 자식 집단을 이용하였다. 재조합 자식 집단은 우리나라 온대 자포니카 초다수 품종인 ‘보람찬’을 모본으로 하고 미국 열대 자포니카 품종인 ‘Pecos’를 부본으로 2013년 인공교배하여 F1 식물체를 양성한 후 육성되었다. F2 분리집단에서 무작위로 200여 개체의 종실 3립씩 채취한 후 F3세대부터 F4세대까지는 단립계통 육종법(SSD; single seed descent method)으로 세대를 진전하였고 이중 187개체를 선발하여 F5세대 187계통을 전개하였다. 이후 F6-F8세대까지 계통당 1개체씩 선발하여 세대진전을 통해 동형접합성이 높아진 187개의 재조합 자식 계통(RIL; recombinant inbred line)을 확보하였다. 본 연구의 시험재료로는 187개 재조합 자식 계통 중 비슷한 농업형질 특성을 나타내는 계통은 제외하고 최종적으로 94개 계통으로 구성된 집단을 모부본의 영문 앞글자를 따 BP_RIL로 명명하여 이용하였다. 2022년 F9세대, 2023년 F10세대 BP_RIL과 모부본을 국립식량과학원 벼 포장에 4월 30일 파종하여 5월 30일에 재식거리 30×15 cm로 2022년에는 주당 1본씩 2열(52주, 26개체/열) 1반복, 2023년에는 주당 1본씩 2열(52주, 26개체/열) 2반복으로 이앙하여 예비선발시험을 수행하였다. 시비량은 완효성 비료인 단한번(성분함량 N-P2O5-K2O-Mg, 18-7-9-2%, (주)조비, Seoul, Korea)을 보비 재배 기준에 따라 기비 시기에 1회 살포하였다. 기타 재배관리는 농촌진흥청 표준 재배법에 준하여 실시하였다.
수량 및 입형 관련 형질 조사
예비선발시험에 공시된 재료의 출수기를 조사하고, 성숙기에 평균이 되는 5개체에 대해서 간장, 수장, 수수를 측정하였다. 성숙기에 3주를 예취하여 등숙률 및 수당립수를 조사하였고 30주를 예취하여 정조중을 측정하였다. 수량성은 측정된 정조중을 30주로 나누어 1주당 수량으로 환산하였다. 수확한 정조 500 g을 제현기(FC2R-Y, Yamamoto Co. Ltd, Yamagata, Japan)로 제영하여 제현율을 측정하였다. 입형 관련 형질을 조사하기 위하여 제현된 현미 중 등숙이 충실이 이루어진 현미를 선별하여 길이, 너비, 두께를 캘리퍼스(Caliper CD-15CP, Mitutoyo Co., Kawasaki, Japan)를 이용하여 5립씩 조사하고 너비에 대한 길이에 비율로 장폭비를 계산하였다. 선별된 현미 100립에 대한 무게를 3반복으로 측정하여 평균 값을 천립중으로 환산하였다.
DNA 추출 및 유전자형 분석
DNA 추출 및 유전자형 분석은 기존에 보고된 방법을 따랐다(
Park et al. 2024b). Genomic DNA 추출은 BioSprint 96 (Qiagen Co., Düren, Germany)을 이용하였다. 샘플을 TissueLyserⅡ (Qiagen Co., Düren, Germany)를 이용하여 마쇄한 후 BioSprint 96 DNA Plant Kit (Qiagen Co., Düren, Germany)를 이용하여 DNA를 추출하였다. BP_RIL과 모부본에 대한 유전자형 분석은 우리나라 자포니카 벼 1,225 SNP, 인디카 벼 1,339 SNP에 대한 정보, 단간 유전자
sd1 내 1 SNP 등 총 2,565개 SNP로 구성된 Target capture sequencing SNP-genotyping platform을 이용하였다(
Cheon et al. 2019,
Lee et al. 2022). Target capture sequencing SNP-genotyping은 (주)인실리코젠(Yongin, Korea) incoFIT Targeted seq panel을 통해 분석되었다. 모부본 게놈에 대한 resequencing은 (주)인실리코젠(Yongin, Korea) incoFIT Genome Sequencing & Analzer package를 통해 분석되었다.
유전자형 분석을 통해 모부본에 다형성을 나타내는 SNP로 762개를 확보하였다. 이들 SNP 중 시험재료의 유전자형에 대한 결측치가 10개 이상인 마커는 제외하고 총 716개 SNP에 대한 유전자형을 수량 및 입형 관련 형질에 대한 QTL 분석에 이용하였다.
유전자지도 작성 및 QTL 분석
유전자지도 작성 및 QTL 분석은 기존에 보고된 방법을 따랐다(
Park et al. 2024b). 유전자 연관지도는 IciMapping Ver. 4.2 (Chinese Academy of Agricultural Sciences, China)의 Kosambi 방법을 이용한 cM (centiMorgan)을 적용하여 작성하였다(
Meng et al. 2015). 모부본에 다형성을 보인 716개 SNP 마커에 대해 12개 염색체의 연관군별로 Grouping하고 Anchor 순서로 Ordering하여 연관군내 마커의 최적순위를 결정하였다.
BP_RIL의 품질 관련 형질에 대한 QTL 분석은 ICIM (Inclusive composite interval mapping) 방법으로 수행하였다. 유의한 QTL을 탐색하기 위한 LOD의 임계값은
p≤0.05 조건으로 1,000회 반복한 permutation test에 의해 결정하였다. QTL 중 2년 연속으로 탐색되었으며, PVE (Phenotypic Variation Explained by the QTL) 값이 20% 이상인 것은 주동 QTL로 규정하였다(
Ashfaq et al. 2023). 탐색된 QTL과 인접한 후보유전자는 Rice Annotation Project Database (RAP-DB,
https://rapdb.dna.affrc.go.jp/, last updated: 11/January/2024)를 이용하여 검색하였다. 후보유전자에 대한 모부본의 염기서열 다형성은 RiceVarMap v2.0 (
https://ricevarmap.ncpgr.cn/, last updated: 2/Feburary/2024)을 이용하여 확인하였다.
통계분석은 R (The R Foundation for Statistical Computing Platform, Version 4.3.3, released: 1/December/2023)을 이용하였다. 수량 및 입형 관련 형질에 대한 평균 등 기술통계는 psych 패키지를 이용하였고, t-test를 이용한 유전자의 대립유전자형에 따른 형질 값의 비교, 분산분석 및 Duncan’s Multiple Range Test (DMRT)를 이용한 대립유전자형 조합에 따른 형질 값의 평균간 비교는 agricolae 패키지를 이용하여 수행하였다. 2년간의 각 형질들의 분포에 대한 히스토그램은 ggplot2, ggpubr, cowplot을 이용하여 작성하였고, 형질들의 표현형 분포 판단은 LaplacesDemon을 이용하였다. 형질들의 상관관계는 corrplot을 이용하여 상관분석을 수행하였고, 형질들의 기여도를 판단하기 위해 lavaan, semPlot, OpenMx, tidyverse, knitr, kableExtra, GGally 패키지를 이용하여 경로계수분석을 수행하였다. Hd1과 Hd6 두 개의 유전자의 상호작용을 알아보기 위하여 Plot2WayAnova 이용하여 이원분산분석을 수행하였다. 출수기 관련 Hd1, Hd6와 입형 관련 qGL2, GW5, qGS10의 대립유전자 및 유전자형 조합에 따른 형질 값을 비교 분석하기 위하여 ggplot2, ggsignif 패키지를 이용하여 vioplin plot을 작성하였고, 입형 관련 형질들의 구조적 관계와 관련 유전자의 대립유전자형 조합에 따른 BP_RIL의 분포를 알아보기 위하여 ggfortify을 이용하여 주성분분석을 실시하였다.
결과 및 고찰
수량 및 입형 관련 형질 표현형 분석
‘보람찬’은 우리나라 온대 자포니카 초다수 품종으로 출수기는 중만생종이며 입형은 중단립종에 단원형 특성을 나타내며 수량성이 높은 것으로 알려져 있으며, ‘Peocs’는 미국 열대 자포니카 품종으로 우리나라 환경조건에서 조생의 출수기를 나타냈으며 중립종에 중원형 입형 특성을 보였고 ‘보람찬’에 비해 간장과 수장이 길고 수수는 적었으며 수당립수가 많고 간이 수량성이 양호한 것으로 보고되었다(
Park et al. 2022). 생태형이 다른 두 품종간 인공교배를 통해 유전적 다양성이 확대된 재조합 자식 집단을 육성하고자 하였고, 동협접합성이 높아진 F
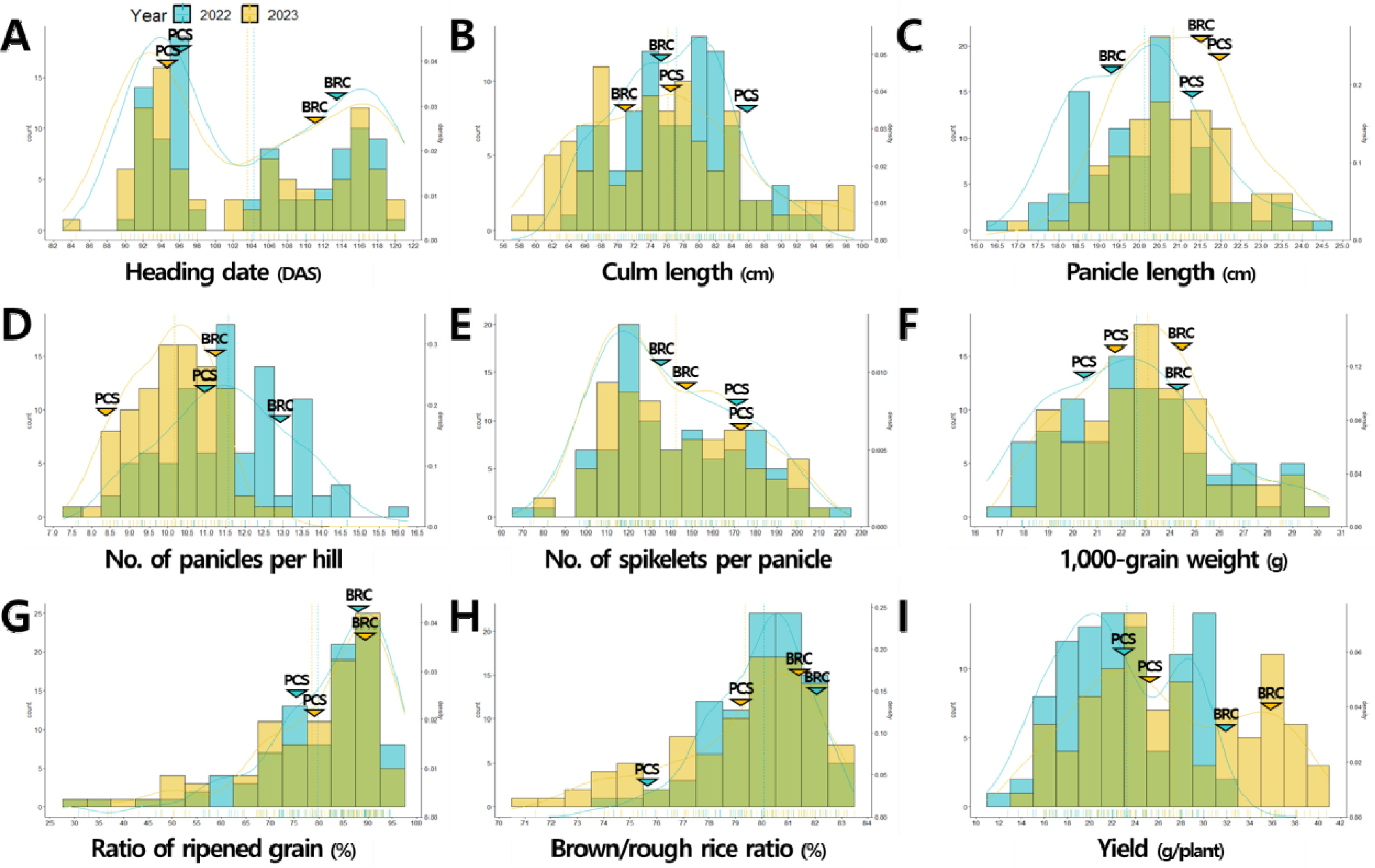
8세대 187개의 RILs 중 비슷한 농업형질 특성을 나타내는 계통은 제외하고 출수기, 입형, 수량 관련 형질에서 표현형 변이가 다양하게 분포하는 94개 RILs을 최종적으로 선발하여 분석에 이용하였다. ‘보람찬/Pecos’ 재조합 자식 집단의 수량 관련 형질QTL 분석을 위하여 모부본과 BP_RIL의 출수기, 간장, 수장, 수수, 수당립수, 천립중, 등숙률, 제현율, 1주 수량을 2022년, 2023년 2년간 조사하였다. 수량 관련 형질들은 대부분 왜도에 의한 치우침은 있으나 평균을 중심으로 연속적인 정규분포를 나타냈고, 2022년 출수기, 간장, 1주 수량, 2023년 출수기, 수당립수, 등숙률, 1주 수량과 같은 일부 형질들은 쌍봉형 분포를 나타냈다(
Fig. 1,
Supplementary Table 1). 여러 유전자가 형질 발현에 영향을 주는 양적형질의 경우 대부분 정규분포를 나타내나 주동 QTL이 작용하는 경우 쌍봉형과 같은 다른 분포를 나타낼 수 있다(
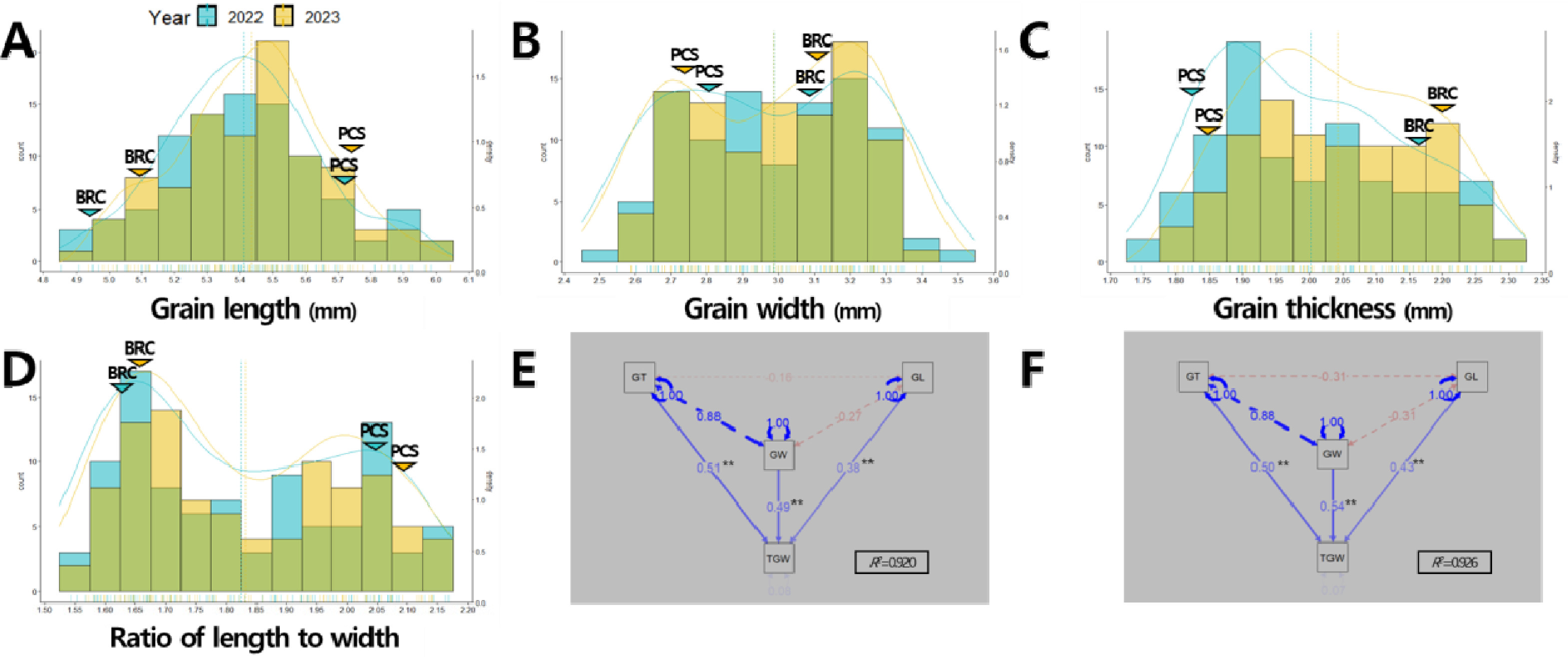
Park et al. 2024b). 2년간 연속적으로 쌍봉형 분포를 나타낸 출수기와 1주 수량의 경우 주동 QTL에 의해서 형질 값이 조절되는 것으로 판단된다. 입형 관련 형질에 대한 표현형 조사 결과, 현미 너비와 장폭비는 2년간 연속적으로 쌍봉형 분포를 나타내 주동 QTL에 의해서 형질 값이 조절되는 것으로 판단되었다(
Figs. 2A-
2D,
Supplementary Table 1). 천립중을 구성하는 입형 관련 형질인 현미 길이, 너비, 두께를 대상으로 한 경로계수분석 결과, 2022년에는 두께(표준화 회귀계수 2022년 0.51), 너비(0.49), 길이(0.38)순, 2023년에는 너비(0.54), 두께(0.50), 길이(0.43) 순으로 천립중에 관여하는 바가 컸다(
Figs. 2E-
2F).
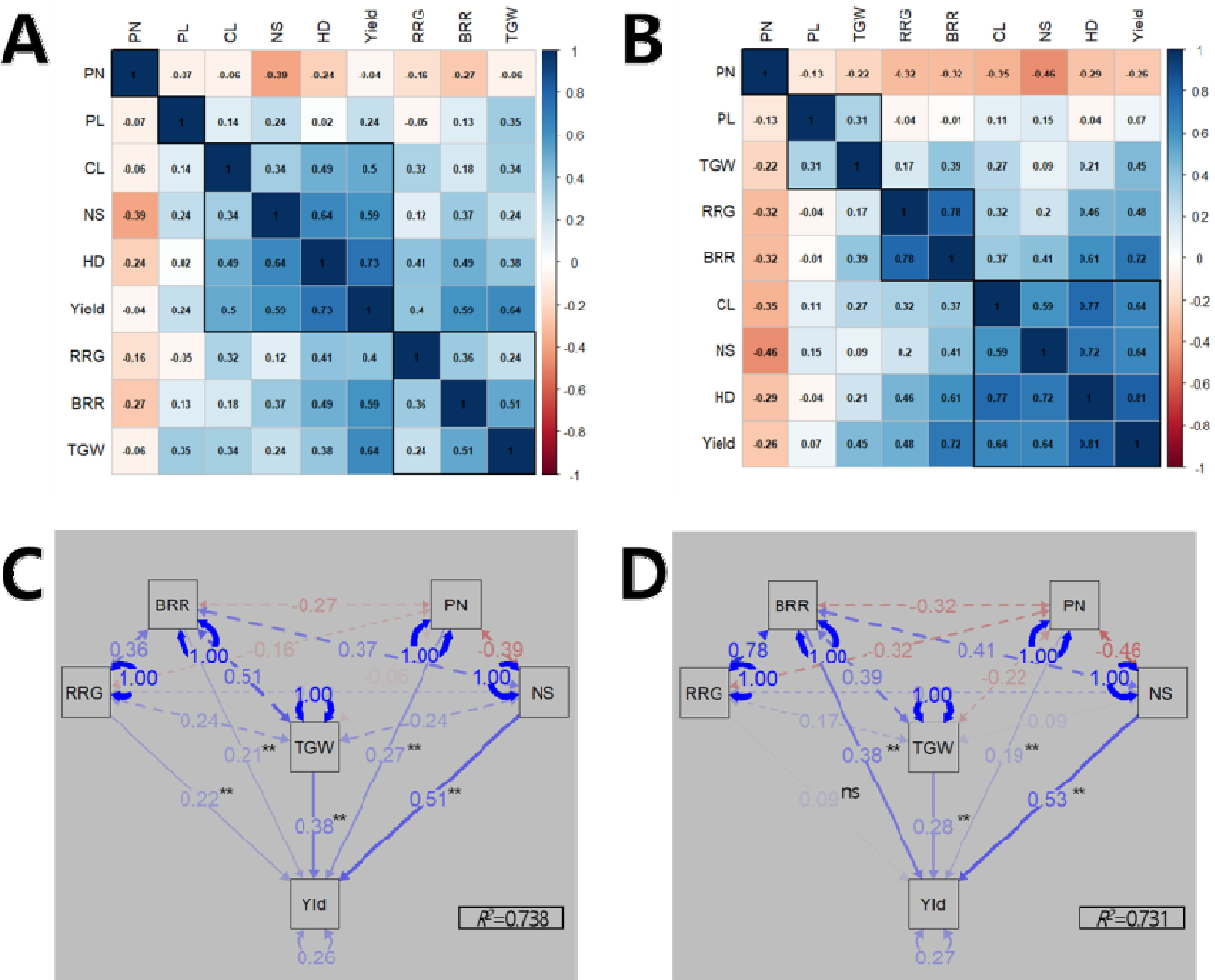
수량 관련 형질들에 대한 관련성을 분석하였다. 상관분석 결과, 출수기는 2년 연속으로 1주 수량, 수당립수, 간장, 제현율, 등숙률, 천립중과 정의 상관관계를 나타낸 반면 수장과는 상관이 적었으며 수수와는 부의 상관을 나타냈다(
Figs. 3A,
3B). 수수는 다른 수량 관련 형질들과 주로 부의 상관을 나타냈는데, 수당립수(상관계수 2022년 -0.39, 2023년 -0.46), 제현율(-0.27, -0.32), 출수기(-0.24, -0.29) 순이었다 수량성에 직접적으로 작용하는 수량구성요소(수수, 수당립수, 천립중, 등숙률, 제현율)들의 1주 수량에 대한 기여율을 분석하기 위하여 경로계수분석을 수행하였다(
Figs. 3C,
3D). 경로계수 분석 결과, 수량구성요소들이 수량성을 설명하는 정도를 나타내는 결정계수(
R2)가 2022년에는 0.738, 2023년에는 0.731를 나타내 70% 이상을 설명할 수 있었다. 2022년에는 통계적으로 유의한 수준에서 수당립수(표준화 회귀계수 0.51), 천립중(0.38), 수수(0.27), 등숙률(0.22), 제현율(0.21) 순으로 1주 수량을 설명하였고, 2023년에는 수당립수(0.53), 제현율(0.38), 천립중(0.28), 수수(0.19) 순으로 수당립수가 2년 연속으로 1주 수량에 기여하는 바가 가장 컸다.
QTL 분석에 이용한 모부본간 다형성을 나타내는 마커는 716개로 염색체 당 마커 수는 35개(9번 염색체)-101개(11번 염색체)로 Mbp당 평균 마커 밀도는 1.3개(6번 염색체)-3.5개(11번 염색체)였다(
Supplementary Table 2). 각 염색체의 물리적 위치(Mbp) 당 마커 수는
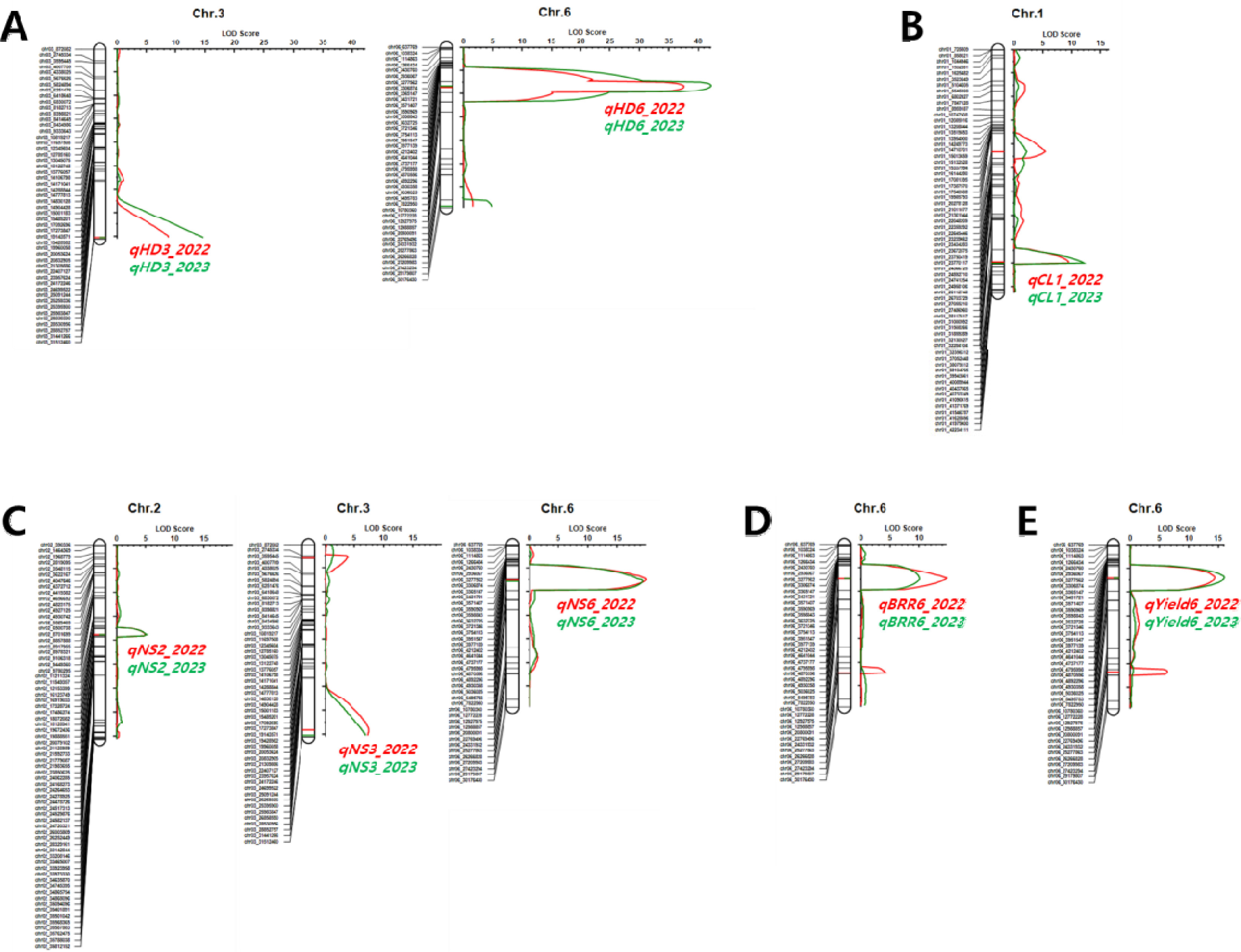
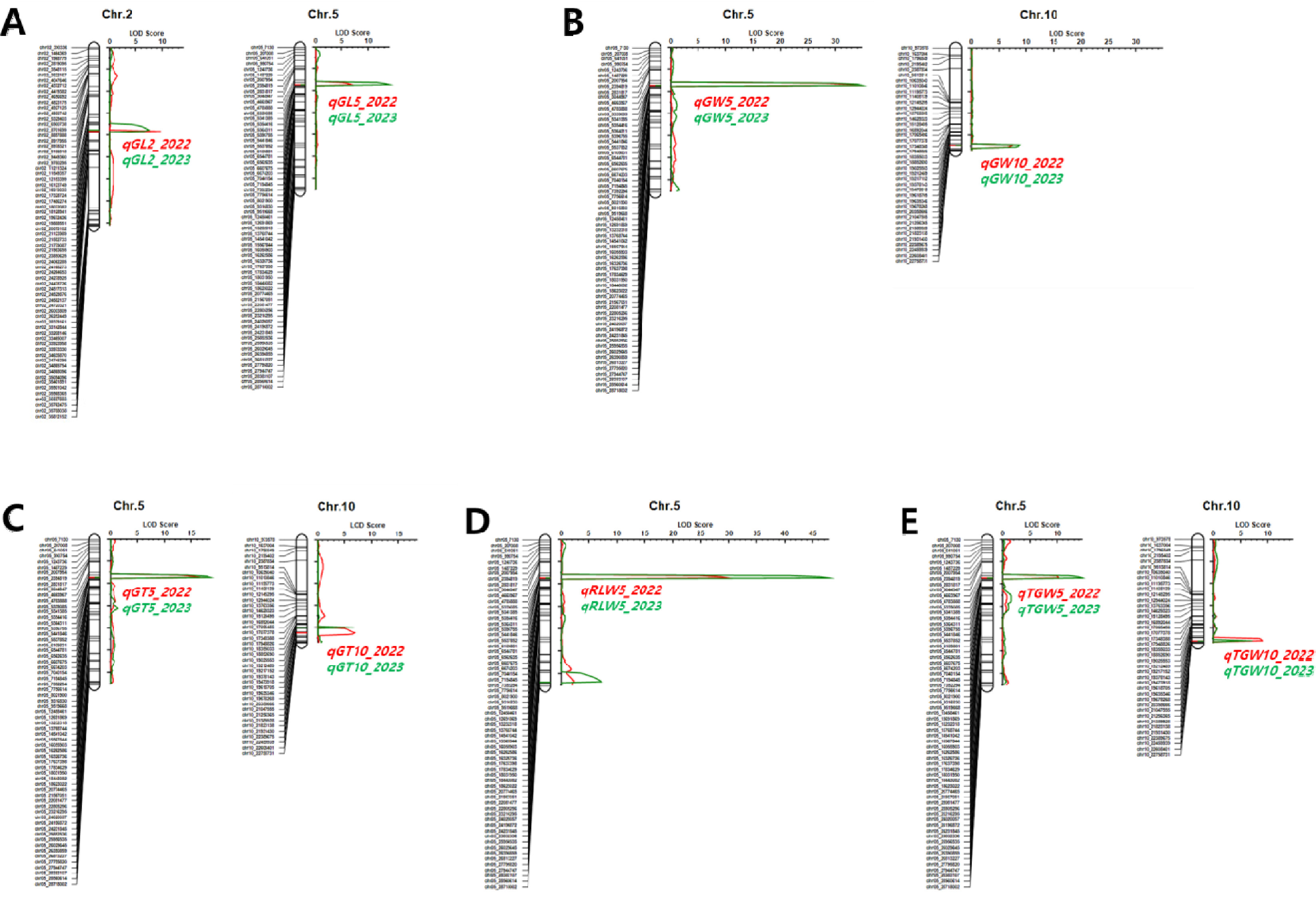
Supplementary Fig. 1과 같다. BP_RIL을 대상으로 2년간 조사된 표현형 값을 이용하여 수량 관련 형질에 대한 QTL을 탐색하였다. 출수기 관련 QTL은 3번과 6번 염색체의 동일한 영역에서 2년 연속으로 탐색되었으며
qHD3과
qHD6로 명명하였다(
Table 1,
Fig. 4A). 3번 염색체에서 탐색된
qHD3는 QTL과 형질 간의 통계적 유의미한 연관성을 나타내는 LOD 값과 QTL이 전체 표현형 변이를 설명할 수 있는 PVE 값이 2022년 8.7, 8.8%였고 2023년은 14.7, 8.4%이었다.
qHD3의 상가적 효과는 -3.54 (2022년), -3.99 (2023년)로 ‘보람찬’의 대립유전자형을 가진 계통은 ‘Pecos’의 대립유전자형을 가진 계통에 비해 출수기가 7일(2022년), 8일(2023년) 가량 빨라졌다. 6번 염색체에서 탐색된
qHD6는 LOD와 PVE 값이 2022년 37.6, 65.4%였고 2023년 42.2, 72.9%로 BP_RIL에 대한 출수기 표현형 변이를 65% 이상 설명할 수 있을 정도의 주동 QTL이었다.
qHD6의 상가적 효과는 9.13 (2022년), 10.83 (2023년)으로 ‘보람찬’의 대립유전자형을 가진 계통은 ‘Pecos’의 대립유전자형을 가진 계통에 비해 출수기가 18일(2022년)에서 22일(2023년) 가량 늦어질 정도로
qHD6 대립유전자 변화에 따른 출수기 변동이 심하였다.
qHD3와
qHD6의 영역에 인접한 출수기 관련 후보유전자를 검색한 결과,
qHD3에서는
Hd6 유전자(
Os03g0762000, chr03:31508813..31514460),
qHD6에서는
Hd1 유전자(
Os06g0275000, chr06:9336376..9338569)를 확인하였다. 6번 염색체 단완에 존재하는
Hd1 유전자는 단일조건에서는 출수를 촉진하고 장일조건에서는 출수를 지연하는 기능을 하여 벼의 재배환경 조건에 따라 상반된 작용을 나타낸다(
Yano et al. 2000). 단일조건인 열대지역에서
Hd1의 기능상실형 대립유전자
hd1-를 보유한 벼의 경우에는 출수가 촉진되는 정상기능형
Hd1+에 비해 출수를 지연시켜 영양생장기간을 늘림으로써 수량성을 확보할 수 있으며, 장일조건인 고위도 지역에서에서는 출수가 지연되는 정상기능형에 비해 출수를 촉진시켜 이른 겨울 전에 수확이 가능하게 함으로써 재배적응성을 높일 수 있다 하였다(
Fujino et al. 2019,
Kim et al. 2018). 우리나라는 고위도에 위치하여 벼의 재배환경은 장일조건이다. 우리나라 293개 벼 품종에 대한
Hd1 대립유전자형 분석 결과, 4개의 정상기능형
Hd1+과 5개의 기능상실형
hd1- 대립유전자형이 존재하는 것으로 보고되었으며,
hd1-을 보유한 품종이
Hd1+을 보유한 품종에 비해 평균 출수일수가 13.7일 단축되는 것으로 나타났다(
Mo et al. 2021). 해당 연구에서 자포니카 중만생종인 ‘보람찬’의 경우 정상기능형
Hd1+ 중 GBZ type (Ginbouzu type)으로 분류되었다(
Mo et al. 2021).
Hd1에 대한 ‘Pecos’의 대립유전자형 분석이 이루어지지 않았으나, 본 연구에서 탐색된 출수기 관련 QTL인
qHD6의 염색체 상 위치(chr06_ 7822950-chr06_10780360)는
Hd1 유전자를 포함하고 있으며, BP_RIL 중 ‘보람찬’의 대립유전자형을 가진 계통들이 ‘Pecos’의 대립유전자형을 가진 계통들에 비해 출수일수가 길어지는 점을 감안할 때
qHD6는
Hd1으로 추정되며, ‘보람찬’은 정상기능형
Hd1+, ‘Pecos’는 기능상실형
hd1-을 보유하고 있으며 향후 ‘Pecos’의 정확한 대립유전자형에 대한 분석이 필요할 것으로 생각된다.
3번 염색체 장완에 존재하는
Hd6 유전자는 단백질 키네이스 CK2의 알파 서브유닛(CK2α)을 암호화하는 벼의 출수기 관련 유전자로 ‘Kasalath’의 정상기능형
Hd6+ 대립유전자는 ‘Nipponbare’의 조기종결 코돈으로 인한 기능상실형
hd6- 대립유전자에 비해 출수일수를 증가시킨다(
Takahashi et al. 2001). RAP-DB를 통한 분석 결과,
Hd6 (
Os03g0762000)는 유전자 내의 31,512,460 bp 위치의 염기서열 차이에 따라 아데닌(A) 염기를 가지고 ‘Kasalath’는 정상기능형
Hd6+, 티민(T) 염기를 보유하고 있는 ‘Nipponbare’는 기능상실형
hd6-을 보유하고 있다. 본 연구에서 탐색된 출수기 관련 QTL인
qHD3의 염색체 상 위치(chr03_28852757-chr03_31441266) 내에
Hd6 유전자가 직접 포함되어 있지는 않으나 인근지역에 위치하고 있으며, BP_RIL의 모부본인 ‘보람찬’과 ‘Pecos’의 벼 표준유전체에 대한 염기서열 분석 결과,
Hd6 내의 31,512,460 bp (functional nucleotide polymorphism; FNP) 위치에서 ‘Pecos’는 ‘Kasalath’와 같은 A 염기를 가지고 있는 반면 ‘보람찬’은 ‘Nipponbare’와 같은 T 염기를 보유하고 있는 것으로 확인되었다(
Supplementary Table 3). 모부본의
Hd6에 대한 염기서열 분석과 BP_RIL 중 ‘Boramchan’의 대립유전자형을 가진 계통들이 ‘Peocs’의 대립유전자형을 가진 계통들에 비해 출수일수가 단축되는 되는 점을 감안할 때
qHD3는
Hd6으로 추정되며, ‘Pecos’는 정상기능형
Hd6+, ‘보람찬’은 기능상실형
hd6-를 보유하고 있는 것으로 판단된다.
간장 관련 QTL은 1번 염색체의 장완에 위치한 마커 chr01_37052448, chr01_38079312사이에서 2년간 연속으로 탐색되었으며
qCL1로 명명하였다(
Table 1,
Fig. 4B). 탐색된 QTL의 LOD 값과 PVE 값은 2022년 9.4, 22.2%였고 2023년은 12.0, 20.6%로 주동 QTL로 판단되었다.
qCL1의 상가적 효과는 -3.21 (2022년), -4.83 (2023년)로 ‘보람찬’의 대립유전자형을 가진 계통은 ‘Pecos’의 대립유전자형을 가진 계통에 비해 간장이 6.4 cm (2022년), 9.7 cm (2023년) 짧아졌다.
qCL1 영역에 인접한 간장 관련 후보유전자를 검색한 결과
SD1 유전자(
Os01g0883800, chr01:38382385.. 38385469)를 확인하였다.
SD1 (
semi-dwarf 1)은 벼의 지베렐린 합성 기작의 거의 마지막 단계를 촉진하는 gibberellin 20 oxidase 2 (GA20ox-2) 암호화하는 반왜성 열성 유전자로 유전자의 기능이 상실된
sd1- 대립유전자형들은 정상기능형
SD1+에 비해 간장이 줄어든다(
Sasaki et al. 2002). 반왜성 열성 유전자
sd1-은 정상 유전자인
SD1+의 염기서열의 결실(deletion), 치환(substitution) 또는 프레임쉬프트 돌연변이 등 다양한 원인에 의한 기능 상실 대립유전자형에 의해 발생하고 많은 수의
sd1 haplotype들이 보고되었다(
Sha et al. 2022). BP_RIL의 모부본인 ‘보람찬’과 ‘Pecos’의 벼 표준유전체에 대한 염기서열 분석 결과 ‘보람찬’은 ‘Reimei’에서 유래한
sd1-r (g.38385083G>C, 3번 엑손 1 bp 치환) 대립유전자형을 보유하고 있고 ‘Pecos’는 정상기능형 대립유전자형인 것으로 확인되어, 간장 관련 탐색된 QTL인
qCL1은
SD1 유전자인 것으로 판단된다(
Supplementary Table 3).
수당립수 관련 QTL은 2번, 3번, 6번 염색체의 동일한 영역에서 2년 연속으로 탐색되었으며
qNS2,
qNS3,
qNS6로 명명하였다(
Table 1,
Fig. 4C). 2번 염색체에서 탐색된
qNS2의 LOD 값과 PVE 값은 2022년 5.0, 6.7%였고 2023년은 5.1, 8.1%이었다.
qNS2의 상가적 효과는 -9.55 (2022년), -9.57 (2023년)로 ‘보람찬’의 대립유전자형을 가진 계통은 ‘Pecos’의 대립유전자형을 가진 계통에 비해 수당립수가 19개씩 줄어들었다.
qNS2가 탐색된 영역에 인접한 수당립수 관련 유전자를 검색(RAP-DB의 Curated genes에서 ‘spikelet number’로 filltering)한 결과, 수당립수와 직접적으로 관련된 유전자는 확인할 수 없었다. 3번 염색체에서 탐색된
qNS3의 LOD 값과 PVE 값은 2022년 7.3, 16.0%였고 2023년은 6.8, 11.9%이었다.
qNS3의 상가적 효과는 -16.08 (2022년), -12.61 (2023년)로 ‘보람찬’의 대립유전자형을 가진 계통은 ‘Pecos’의 대립유전자형을 가진 계통에 비해 수당립수가 32개(2022년), 25개(2023년) 줄어들었다. 6번 염색체에서 탐색된
qNS6의 LOD 값과 PVE 값은 2022년 19.9, 40.4%, 2023년 19.3, 44.8%로 주동 QTL로 판단되었다.
qNS6의 상가적 효과는 23.42 (2022년), 22.41 (2023년)로 ‘보람찬’의 대립유전자형을 가진 계통은 ‘Pecos’의 대립유전자형을 가진 계통에 비해 수당립수가 47개(2022년), 45개(2023년) 늘어났다.
qNS3와
qNS6가 탐색된 영역은 출수기 관련 QTL
qHD3,
qHD6와 동일한 위치이며 후보 유전자로
Hd6와
Hd1이 추정되었다(
Table 1). 하나의 형질에 관여하는 QTL에 의해서 다른 형질들도 영향을 받는 다면발현 효과들이 보고되었다(
Seo et al. 2014). 출수기 관련 유전자는 수당립수와 밀접한 관련성을 가지고 있는 것으로 알려져 있어(
Lu et al. 2022,
Sun et al. 2023),
Hd6와
Hd1의 다면발현으로 인하여 수당립수가 영향을 받아 해당 영억에서
qNS3와
qNS6가 탐색된 것으로 판단된다.
제현율과 1주 수량 관련 QTL인
qBBR6과
qYield6은 표현형에 대한 설명정도가 20%가 넘는 주동 QTL로 출수기 관련 QTL인
qHD6와 동일한 위치에서 탐색되었다(
Table 1,
Figs. 4D, 4E).
qHD6의 후보유전자인
Hd1은 출수기 뿐만 아니라 수량 관련 형질에 대해서 영향을 미치는 것으로 알려져 있어
Hd1의 기능에 의해서 제현율과 1주 수량이 조절된 것으로 판단된다(
Lee et al. 2024,
Zhang et al. 2012). 상가적 효과에 대한 분석결과 ‘보람찬’의 대립유전자형을 가진 계통이 ‘Pecos’의 대립유전자형을 가진 계통에 비해 제현율이 높고 1주 수량이 증가하였다.
Hd1에 대한 ‘보람찬’의 대립유전자형은 정상기능형
Hd1+로 ‘Pecos’의 기능상실형
hd1-에 비해서 출수일수가 증가함으로써 우리나라 환경조건에서 고온에 의한 등숙조건을 회피함으로써 등숙이 양호하여 제현율이 높아진 것으로 판단된다. 또한 정상기능형
Hd1+ 대립유전자형은 제현율 향상 뿐만 아니라 수당립수의 증가에도 영향을 미쳤는데 이에 따라 1주 수량이 증가한 것으로 판단된다. 출수기 관련
Hd1 유전자는 출수기 뿐만 아니라 수당립수, 제현율, 수량성 등 주요 수량 관련 형질 변이에도 영향을 미치는 것으로 확인되어 하나의 유전자가 여러 가지 형질에 영향을 미치는 현상인 다면발현을 나타냈다.
BP_RIL을 대상으로 2년간 조사된 표현형 값을 이용하여 입형 관련 형질에 대한 QTL을 탐색하였다. 현미 길이, 너비, 두께, 장폭비, 천립중 관련 QTL이 2번, 5번, 10번 염색체에서 2년 연속으로 탐색되었다(
Table 2,
Fig. 5). 5번 염색체에서 탐색된 QTL은 현미 길이(
qGL5), 너비(
qGW5), 두께(
qGT5), 장폭비(
qRLW5) 및 천립중(
qTGW5) 등 입형 관련 형질 모두 관여하며 PVE가 21.1-76.2%로 입형 관련 형질 변이의 주동 QTL로 확인되었다. ‘보람찬’의 대립유전자형을 가진 계통은 ‘Pecos’의 대립유전자형을 가진 계통에 비해 현미 길이는 짧아지고 너비와 두께는 증가하여 장폭비는 줄어들었고 천립중은 증가하였다. 해당 QTL 영역(chr05_4783888-chr05_5339085)에 인접한 입형 관련 유전자를 검색한 결과,
qSW5/GW5 (
Os05g0187500, chr05:5365122..5366701)를 확인하였다.
qSW5/GW5는 종실의 너비와 무게에 관여하는 유전자로 기존 연구결과에서 ‘보람찬’은 ‘Nipponbare’와 같이 기능상실형 대립유전자인
qsw5_N을 보유하고 ‘Pecos’는 ‘Kasalath’와 같은 정상기능형
qSW5을 가지고 있는 것으로 보고되어(
Park et al. 2022), 5번 염색체에서 탐색된 QTL의 후보유전자는
qSW5/GW5로 판단된다.
2번 염색체에서 탐색된 QTL은 현미 길이(
qGL2)에 대해서만 영향을 미쳤고, 10번 염색체에서 탐색된 QTL은 너비(
qGW10), 두께(
qGT10), 천립중(
qTGW10)에 관여하는 것으로 나타났다.
qGL2가 탐색된 영역 근처에서 현미 길이 관련 유전자를 검색(RAP-DB의 Curated genes에서 ‘grain length’로 filltering)한 결과,
OML4 (
Os02g0517531, chr02:18769958.. 18777995)를 확인하였다.
OML4 (
MEI2-LIKE PROTEIN4)는 종실크기에 관여하는
LARGE1과 동일유전자로 글리코겐 합성효소 키나제2 (
GSK2)에 의해 인산화되어 벼의 종실 크기와 무게를 부정적으로 조절하게 되는데
OML4의 기능이 상실되면 종실의 크기가 커지고 무게가 증가하는 것으로 보고되었다(
Lyu et al. 2020).
OML4 내 모부본인 ‘보람찬’과 ‘Pecos’의 벼 표준유전체에 대한 염기서열 분석 결과, ‘보람찬’은 변이가 탐색되지 않았고, ‘Pecos’는 2개의 SNP (vg0218771599, intron_variant ; MODIFIER, vg0218774768, synonymous_variant ; p.Gly556Gly; LOW)와 3개의 INDEL (vg0218770045, vg0218770186, vg0218776158, intron_variant; MODIFIER)이 탐색되었다. 물리적 위치의 인접성, 현미 길이에 관여하는 기능적 유사성과 염기서열 변이의 탐색 등을 고려할 때
qGL2의 후보유전자로
OML4이 추정되나 정확한 확인을 위해서는 추가 연구가 필요할 것으로 판단된다.
10번 염색체에서 탐색된 QTL은
qGW10 (chr10_21256365-chr10_21389928),
qGT10_2022 (chr10_20358666- chr10_21047555),
qGT10_2023 (chr10_19473918-chr10_19618705),
qTGW10 (chr10_21823138-chr10_21931430)로 10번 염색체 장완 하단의 19.4-21.9 Mbp에 위치하고 있었다. 해당 영역에 인접한 입형 관련 유전자를 검색(RAP-DB의 Curated genes에서 ‘grain size’로 filtering)한 결과,
GW10 (
Os10g0515400, chr10:19863346..19865997),
GS10 (
Os10g0522601, chr10:20263310..20267568),
OsCPq10 (
Os10g0525200, chr10:20426804..20430567)를 확인하였다.
GW10은 cytochrome P450 subfamily 단백질을 암호화하며 브라시노스테로이드(BR) 경로에 관여해 종실크기와 수당립수에 영향을 미치며,
OsCPq10 또한 cytochrome P450 family 단백질을 암호화며 종실 크기에 영향을 미치는 것으로 보고되었다(
Park et al. 2023,
Zhan et al. 2021).
GS10은 6개의 armadillo 반복 도메인을 가진 단백질을 암호화하며 BR 신호 전달 경로에서 음성 조절자로 작용하여 벼의 종실 크기를 조절하는 역할을 하는데,
GS10은
OsGSK2 (
Glycogen Synthase Kinase 2)와 후성적(epistatic) 관계를 가지는 것으로 보고되었다(
Chen et al. 2023).
OsGSK2는 BR 신호 전달 경로의 음성 조절자로 작용하는
GSK3/SHAGGY-like kinase로
OsBZR1과
DLT와 같은 전사인자를 인산화하여 그 활성을 억제함으로써 BR 신호를 조절한다(
Liu et al. 2017). 2번 염색체에서 탐색된
OML4/LARGE1도
OsGSK2에 의해 인산화되어 종실 크기를 음성적으로 조절하고, 5번 염색체에서 탐색된
qSW5/GW5는
OsGSK2의 활성을 억제함으로써 BR 신호를 양성적으로 조절하는 등 2, 5, 10번 염색체에서 탐색된 입형 관련 QTL의 주요 후보유전자들은
OsGSK2와 상호작용을 하여 BR 신호 전달 경로에 관여함으로써 입형 관련 형질 변이에 영향을 미치는 것으로 판단된다(
Chen et al. 2023,
Liu et al. 2017,
Lyu et al. 2020). 10번 염색체에서 탐색된 QTL의 후보유전자를 추정하기 위해 BP_RIL의 모부본인 ‘보람찬’과 ‘Pecos’의 벼 표준유전체에 대한 염기서열 분석 결과, ‘보람찬’은
GW10,
GS10,
OsCPq10 모두에서 변이가 탐색되지 않았고, ‘Pecos’는
GW10에서 20개의 SNP (vg1019864829 등 10개 missense_variant; MODERATE, vg1019864591 등 10개 synonymous_variant; LOW)와 1개의 INDEL (vg1019865301, inframe_insertion ; p.Ala175dup; MODERATE) 변이가 탐색되었고
GS10에서 1개의 SNP (vg1020263841, upstream_gene_variant ; 1380.0bp to feature; MODIFIER),
OsCPq10에서 4개의 SNP (vg1020427937 등 3개 upstream_gene_variant; MODIFIER, vg1020430474, synonymous_variant ; p.Asp20Asp; LOW)가 탐색되었다. 후보유전자에 대한 탐색된 변이의 개수 및 변이가 단백질 기능에 미치는 영향 등을 감안했을 때 10번 염색체에서 탐색된 입형 관련 QTL의 후보유전자는
GS10,
OsCPq10에 비해
GW10일 가능성이 높은 것으로 여겨지나 탐색된 QTL의 위치가 형질과 연차에 따라서 변동되는 점 등을 고려할 때 정확한 후보유전자의 확인을 위해서는 추가 연구가 필요할 것으로 판단된다. 해당 QTL은 현미 너비, 두께, 천립중 등 입형 크기에 관여하여
qGS10 (
QTL for grain size on chromosome 10)으로 명명하고 추가 분석을 수행하였다.
수량 관련 형질 QTL의 후보유전자인
Hd1,
Hd6,
SD1가 수량 관련 형질에 미치는 영향을 분석하였다. 출수기 관련 유전자
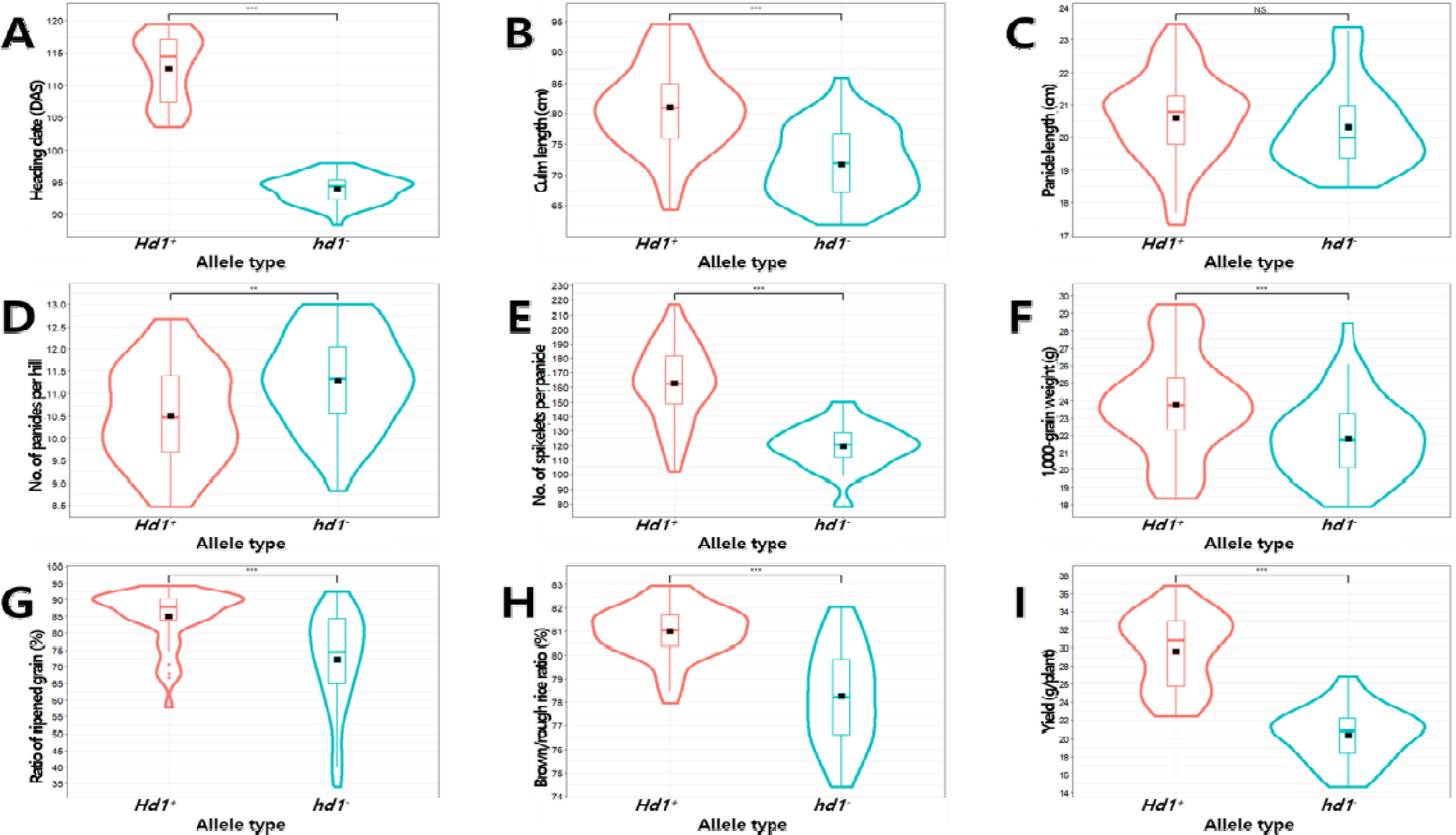
Hd1은 수장을 제외하고 출수기, 간장, 수수, 수당립수, 천립중, 등숙률, 제현율, 1주 수량에 영향을 미치는 것으로 나타났다(
Table 3,
Fig. 6). 정상기능형 대립유전자형
Hd1+은 기능상실형
hd1-에 비해 출수일수가 19일 늦어졌으며 간장이 9 cm 길었고 수수가 0.8개 감소하였으며 수당립수는 44개 증가하였고 천립중이 2.0g 무거웠으며 등숙률과 제현율이 각각 12.8%, 2.8% 높아졌으며 1주 수량이 8.2 g 증가하였다.
Hd1은 출수기 이외의 다른 형질에 영향을 미치는 등 다면발현 현상이 보고되었다. 장일조건인 우리나라 환경조건에서 국내 육성품종들에 대한 분석에서
Hd1+ 대립유전자형은
hd1- 비해 출수일수가 늦고 간장이 길었으며, 단일조건인 저위도 지역에서는 일장에 둔감한
hd1- 대립유전자형이
Hd1+에 비해서 기본영양생장성(BVP)이 높아 출수일수가 늘어날 뿐만 아니라 간장이 길어지고 수당립수가 증가하며 수량성이 높아지는 등 출수기 이외에 수량 관련 형질 변이에도 영향을 미치는 것으로 나타났다(
Mo et al. 2021,
Zhang et al. 2012). 본 연구에서 대부분의 수량 관련 형질들이
Hd1 대립유전자형에서 표현형 값이 증가하는 등 생육기간이 길어짐에 따라서 형질 값이 증가하는 양상을 보였으나 수수에서는 반대의 경향을 나타내
Hd1과 수수와의 다면발현 양상에 대해서는 추가 분석이 필요할 것으로 판단된다.
또 다른 출수기 관련 유전자
Hd6는 대립유전자형간 수장과 등숙률에서만 통계적으로 유의한 차이가 있는 것으로 나타났다(
Table 3).
Hd6는 QTL 분석을 통해 출수기와 수당립수에 대한 후보유전자로 탐색되었으나 전체 집단의 표현형 변이를 설명하는데 있어서는
Hd6의 대립유전자형에 따른 출수기와 수당립수의 차이는 없는 것으로 나타났다(
Tables 1,
3). 이러한 이유로는 출수기 관련 유전자인
Hd6와
Hd1과의 상호작용(상위성) 효과가 존재하거나
Hd1이 출수기와 수당립수의 표현형 변이에 미치는 PVE (출수기 65.4% (2022년), 72.9% (2023년), 수당립수 40.4% (2022년), 44.8% (2023년))가
Hd6 (출수기 8.8%, 8.4%, 수당립수 16.0%, 11.9%)에 비해 높기 때문에
Hd1의 영향에 의해서
Hd6가 출수기 등 수량 관련 표현형 변이에 미치는 영향이 가려짐에 따라 대립유전자형에 따른 형질 값의 차이가 나지 않은 것으로 추정된다. 출수기 변이에 큰 영향을 미쳐 주동 유전자로 판단되는
Hd1과 이에 비해 미동 유전자인
Hd6가 수량 관련 형질 변이에 미치는 영향을 ANOVA를 통해 분석한 결과,
Hd1의 F 값이 1003.5로
Hd6 (81.0)에 비해 컸으며 이외에 간장, 수수, 수당립수, 천립중, 등숙률, 정현비율, 1주 수량에 미치는 영향도 큰 것으로 나타났다(
Supplementary Table 4). 간장 관련 유전자
SD1은 ‘보람찬’의 단간 대립유전자형
sd1-을 보유한 계통들이 ‘Pecos’의
SD1+을 보유한 계통들이 비해 간장이 평균 7 cm 가량 짧아졌으며, 이외의 다른 형질들의 변이에는 영향을 미치지 않는 것으로 나타났다(
Table 3).
주동 유전자에 의한 미동 유전자의 작용이 가려지는(masking) 현상을 파악하기 위해서
Hd1과
Hd6의 대립유전자형 조합에 따른 분석을 통해 주동 유전자인
Hd1의 대립유전자형을 고정시키고
Hd6의 대립유전자형에 따른 수량 관련 형질의 차이를 추가 분석하였다. 출수기 관련 유전자인
Hd1은 출수기, 수당립수, 정현비율, 1주 수량의 후보유전자로 추정되었으며 대립유전자형에 따라 해당 형질 값이 통계적으로 유의미한 차이를 보여 출수기 이외의 다른 형질 변이에도 영향을 미치는 다면발현 현상을 나타냈다(
Tables 1,
3). 반면, 또 다른 출수기 관련 유전자인
Hd6는 출수기와 수당립수의 후보유전자로 추정되었으나 대립유전자형에 따른 형질 값의 통계적 차이는 관찰되지 않았다. 출수기 관련 유전자 간 상호작용 효과를 분석하기 위해
Hd1과
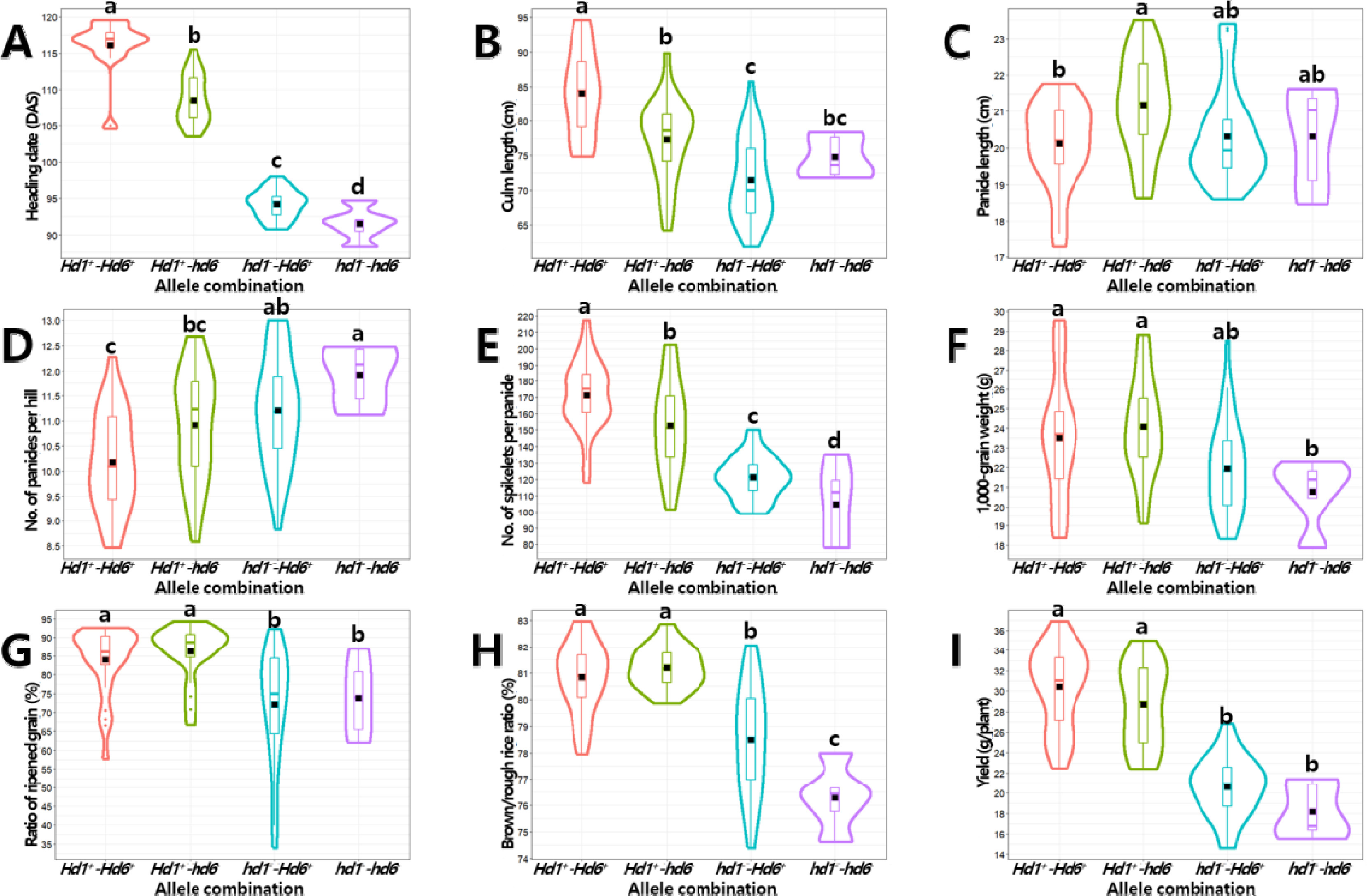
Hd6 대립유전자형 조합에 따른 수량 관련 형질 값을 비교 분석하였다(
Table 4,
Fig. 7). BP_RIL 중 모본인 ‘보람찬’과 같은 대립유전자형 조합인
Hd1+-
hd6-을 가지는 계통이 23개, 부본인 ‘Pecos’와 같은 대립유전자형 조합인
hd1--Hd6+을 가지는 계통이 39개로 양친형의 수는 62개였으며, 재조합형인
Hd1+-Hd6+ (27계통)와
hd1--hd6- (5계통) 대립유전자형 조합이 32개로 재조합 빈도는 34.0%를 나타냈다.
Hd1과
Hd6가 서로 다른 염색체에 존재함에도 재조합 빈도가 낮았던 것은
hd1-을 보유한 조합 중
hd1--Hd6+ (Pecos형, 39계통)과
hd1--hd6- (5계통)이 선발과정에서 편이가 발생했기 때문으로 판단된다. 전체 187개의 재조합 자식 계통 중 94개의 BP_RIL을 선정하는 과정에서 형질 특성이 일부 열악했던
hd1--hd6- 계통들에 선발압이 작용했을 것으로 추정된다. 출수기는 정상기능형 대립유전자들로 구성된
Hd1+-Hd6+ 조합에서 가장 길었으며(출수일수 116일), 다음으로
Hd1+-hd6- (109일),
hd1--Hd6+ (94일), 기능상실형 대립유자들로 구성된
hd1--hd6- (92일) 순으로 나타났다(
Table 4,
Fig. 7A). 수당립수 역시 출수기와 유사한 경향을 보여
Hd1+-Hd6+ (171개),
Hd1+-hd6- (153개),
hd1--Hd6+ (121개),
hd1--hd6- (105개) 순으로 형질 값이 컸다. 출수기와 수당립수의 후보유전자로 추정된
Hd6는 전체 집단의 표현형 변이를 설명하는데 있어서는 대립유전자형간 형질 값에서 통계적 차이가 나타나지 않았다(
Table 3). 그러나
Hd1+의 정상기능형을 보유한 조합간 비교(
Hd1+-Hd6+와
Hd1+-hd6-)에서는 정상기능형
Hd6+를 가진 계통이 기능상실형
hd6-에 비해 출수일수가 평균 7일 더 길고 수당립수가 18개 더 많은 것으로 나타났다(
Table 4,
Figs. 4A,
4E).
hd1- 기능상실형을 보유한 조합 간 비교(
hd1--Hd6+와
hd1--hd6+)에서도
Hd6+를 보유한 계통이
hd6-에 비해 출수일수가 평균 2일 더 길고 수당립수가 16개 더 많은 것으로 확인되었다(
Table 4,
Figs. 4A,
4E). 이원분산분석 결과, 출수기 표현형 변이에
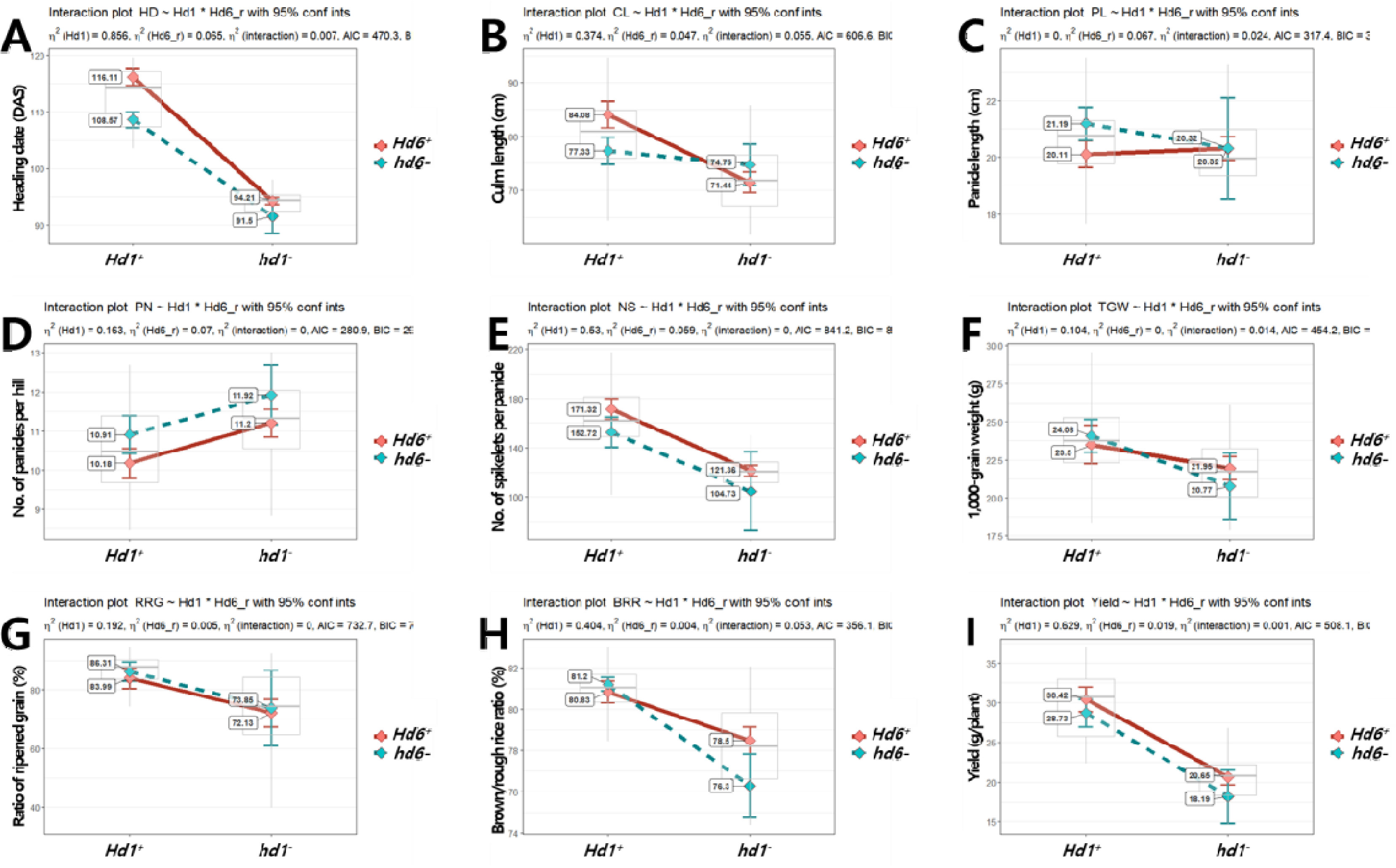
Hd1이 미치는 영향(부분 에타제곱(
η2): 0.856)은
Hd6 (0.065)에 비해 매우 컸으며 상호작용효과(0.007)는 낮아
Hd1이 출수기 변이에 주동 유전자로 작용한 것으로 나타났다(
Fig. 8). 수당립수에서도
Hd1의 영향력(부분 에타제곱(
η2): 0.530)이
Hd6 (0.059)에 비해 컸으며 상호작용효과(0)가 없는 것으로 나타나 수당립수 변이에서도 주동 유전자는
Hd1이고,
Hd6는 출수기와 수당립수 형질 변이에 미동 유전자로 작용한 것으로 판단된다. 출수기와 수당립수 변이에 미동 유전자로 관여하는
Hd6는 주동 유전자인
Hd1 대립유전자형이 고정된 경우 대립유전자형간 형질 값에서 통계적 차이가 존재하나 전체 집단의 표현형 변이에 있어서는 그 효과가 확인되지 않는 등 주동 유전자에 의해 미동 유전자의 작용이 가려지는(masking) 현상이 발생하였다.
출수기와 수당립수 등 대부분의 수량 관련 형질들은 영양생장기간이 증가하게 되는
Hd1와
Hd6 정상기능형 대립유전자가 집적될수록 형질 값이 증가하는 경향을 보였으나, 수수의 경우 반대 경향을 나타냈다(
Figs. 7,
8). 수당립수와 수수는 종실크기와 함께 동일한 자원을 사용하는 sink (동화산물이 이동하여 저장되는 장소)로 작용하므로 하나의 형질이 증가하면 다른 형질은 감소하는 상충 관계(trade-off)를 나타내는 경우가 많다(
Fan & Li 2019). 본 연구에서 수당립수와 수수는 부의 상관관계를 나타냈으며(
Fig. 3), 다른 많은 연구결과에서도 경쟁적 관계를 나타냈다(
Park et al. 2015,
Park et al. 2020,
Park et al. 2023).
Hd1과
Hd6의 정상기능형 대립유전자형의 집적 시 수수가 줄어든 원인으로는 출수 지연으로 축적된 에너지가 유효분얼 개념인 수수의 증가보다는 영화 형성에 따른 수당립수 증가로의 배분이 많이 이루어짐에 따라 발생한 것으로 추정되나 정확한 원인에 대해서는 추가분석이 필요할 것으로 생각된다.
입형 관련 QTL과 후보유전자인
qGL2,
qSW5/GW5 (이후
GW5 표기),
qGS10이 입형과 수량 관련 형질에 미치는 영향을 분석하였다.
qGL2는 ‘Pecos’의 대립유전자형인
qGL2P를 보유한 계통(n=43)이 ‘보람찬’의 대립유전자형
qGL2B를 보유한 계통(n=51)에 비해 통계적으로 현미 길이가 길고 천립중이 무거웠다(
Fig. 9).
qGL2는 제현율을 제외한 수량 관련 형질에 영향을 미쳤는데,
qGL2P 대립유전자형이
qGL2B에 비해 출수가 지연되며 간장과 수장이 길어지고 수당립수는 증가하면서 수수는 감소하였고 등숙률과 천립중이 높고 1주 수량이 높은 것으로 나타났다(
Table 5). 본 연구에서
qGL2는 수당립수 QTL인
qNS2와 동일영역에서 탐색되었으며 후보 유전자로
OML4이 추정되었다(
Table 2).
OML4는 종실크기에 관여하는
LARGE1, 출수기에 관여하는
EHD6와 동일 유전자로 확인되었다(
Cui et al. 2024,
Lyu et al. 2020).
qGL2 영역은 이전 보고에서 종실크기와 출수기에 대한 유전자로 동정되었고, 본 연구결과에서 현미 길이에 대한 QTL로 탐색되었으며 입형 뿐만 아니라 수당립수 등 수량 관련 형질에도 다양하게 영향을 미치는 것으로 확인되었다. 미국품종 ‘Pecos’에서 유래한
qGL2P 대립유전자형은 현미 길이가 길어지고 천립중이 증가하였으며 수당립수가 많아짐에도 높은 등숙률을 나타내면서 1주 수량이 증가하는 것으로 나타나 우리나라 자포니카 벼 품종으로 도입하여 활용할 가치가 있는 유용인자로 판단된다.
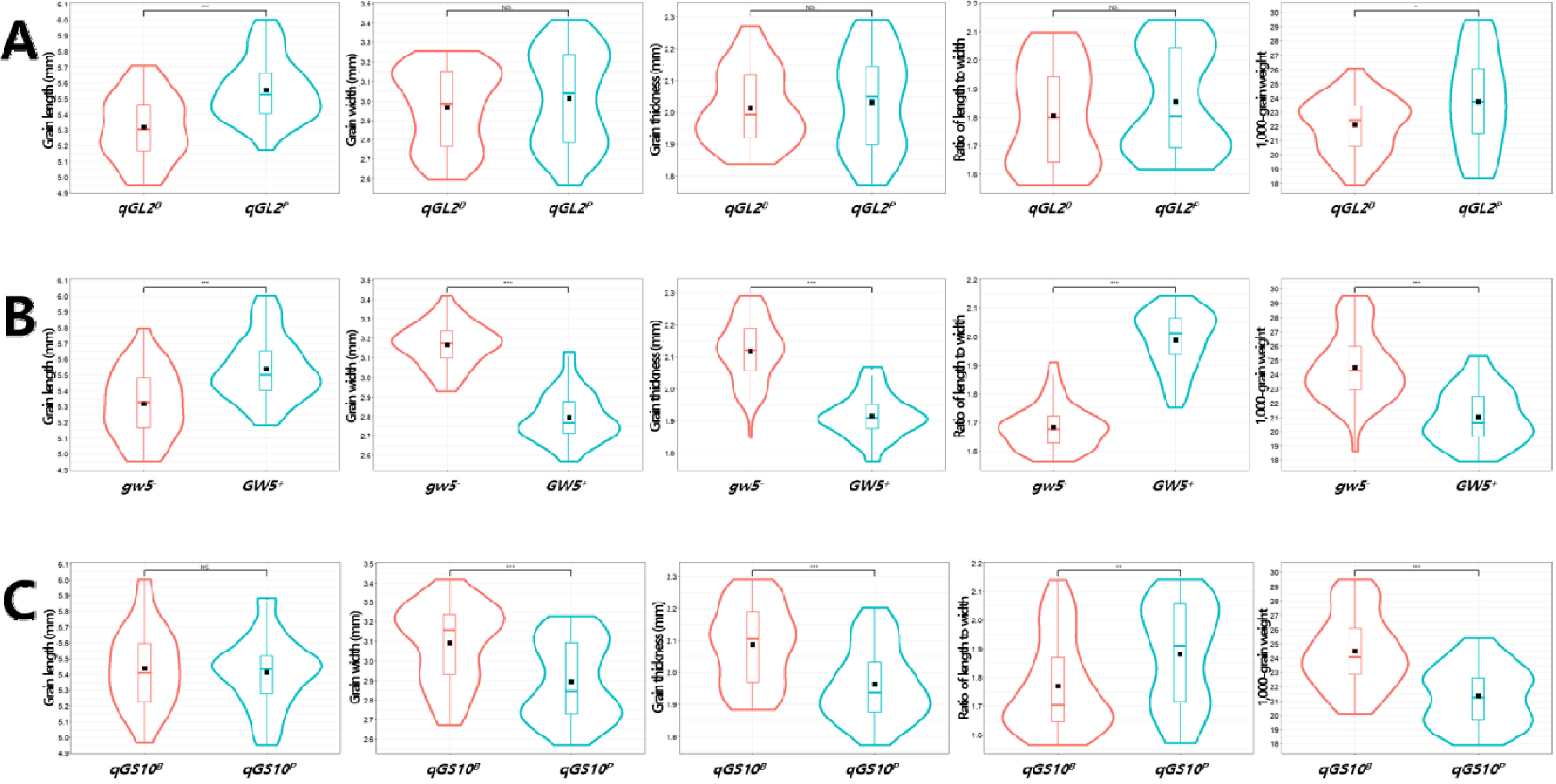
GW5는 ‘보람찬’의 대립유전자형인
gw5-를 보유한 계통(n=49)이 ‘Pecos’의 대립유전자형
GW5+를 보유한 계통(n=45)에 비해 통계적으로 현미 너비와 두께가 길고 천립중이 무거웠으며 길이가 짧고 장폭비가 작았다(
Fig. 9).
GW5는 출수기, 천립중, 등숙률, 정현비율 및 1주 수량에 영향을 미쳤는데,
gw5- 대립유전자형이
GW5+에 비해 출수가 지연되며 천립중, 등숙률 정현비율, 1주 수량이 높았다(
Table 5). 우리나라 대부분의 자포니카 벼 품종들은 ‘보람찬’과 같이
gw5- 대립유전자형을 보유하고 있고 미국 품종들은 ‘Pecos’와 같이
GW5+를 나타내는 것으로 보고되었는데(
Lee et al. 2020,
Park et al. 2022), 본 연구결과에서
gw5- 대립유전자형이
GW5+에 비해 수량성을 증가하는 방향으로 작용하였다. 이를 통해 우리나라 자포니카 벼 품종 육성에 있어서 국내 환경조건에서 적응성이 높은
gw5- 대립유전자형에 대한 선발압이 작용하여 대부분의 품종들이
gw5-를 보유하고 있는 것으로 추정된다.
qGS10는 ‘보람찬’의 대립유전자형인
qGS10B를 보유한 계통(n=45)이 ‘Pecos’의 대립유전자형
qGS10P를 보유한 계통(n=49)에 비해 통계적으로 현미 너비와 두께가 길고 천립중이 무거웠으며 장폭비가 작았다(
Fig. 9).
qGS10은 출수기, 간장, 수장, 수당립수, 천립중, 1주 수량에 영향을 미쳤는데,
qGS10B 대립유전자형이
qGS10P에 비해 출수가 지연되며 간장과 수장이 길고 수당립수가 많았으며 천립중과 1주 수량이 높았다(
Table 5). 우리나라 ‘보람찬’이 보유한
qGS10B 대립유전자형은 천립중이 무거우면서도 수당립수가 많은 특성을 나타냈다. 벼의 수량구성요소 중 종실 크기와 립수는 한 쪽이 증가하면 다른 한쪽이 감소하는 상충 관계(trade-off)를 주로 나타는 것으로 알려져 있다(
Fan & Li 2019). 우리나라 자포니카 벼 입형 다양화 육종소재와 ‘참동진/영호진미’ 재조합 자식 집단의 수량구성요소들간 상관분석에서도 수당립수와 천립중은 부의 상관관계를 나타냈으며 천립중이 증가한 자원의 경우 수당립수가 감소하여 수량성을 향상에 제한요인으로 작용하였다(
Park et al. 2017,
Park et al. 2024b). 이에 반해 본 연구에서 확인된 ‘보람찬’이 보유한
qGS10B 대립유전자형의 경우 천립중이 무거워지면서 수당립수도 함께 증가하여 수량성 향상을 위한 유용인자로 판단되었다. 하지만 통계적으로 유의미하지는 않지만
qGS10P에 비해
qGS10B가 등숙률이 감소하는 것으로 나타나 다른 입형 관련 유전자와의 상호작용에 따른 수량성에 미치는 영향도 고려해 보야야 할 것으로 생각된다.
qGL2, GW5, qGS10 대립유전자형 조합이 입형 및 수량 관련 형질에 미치는 영향
입형 관련 QTL과 후보유전자인
qGL2,
GW5,
qGS10의 대립유전자형 조합이 입형 관련 형질에 미치는 영향과 유전자들의 상호작용 효과를 분석하였다. ANOVA 분석 결과, 현미 길이에 대해서는
qGL2와
GW5가 비슷한 수준으로 영향을 미쳤으며 너비, 두께, 장폭비에 대해서는
GW5가 가장 큰 영향을 미쳤고 천립중에 대해서는
GW5,
qGS10,
qGL2 순으로 관여하는 것으로 나타났고, 대립유전자간 상호작용 효과는 현미 길이에 대해서
qGL2*
qGS10, 천립중에 대해서
qGL2*
GW5가 일부 관여하였다(
Supplementary Table 5). BP_RIL 내에는 세 개의 유전자로 작성 가능한 8개의 대립유전자형 조합이 모두 존재하는 것으로 나타났다(
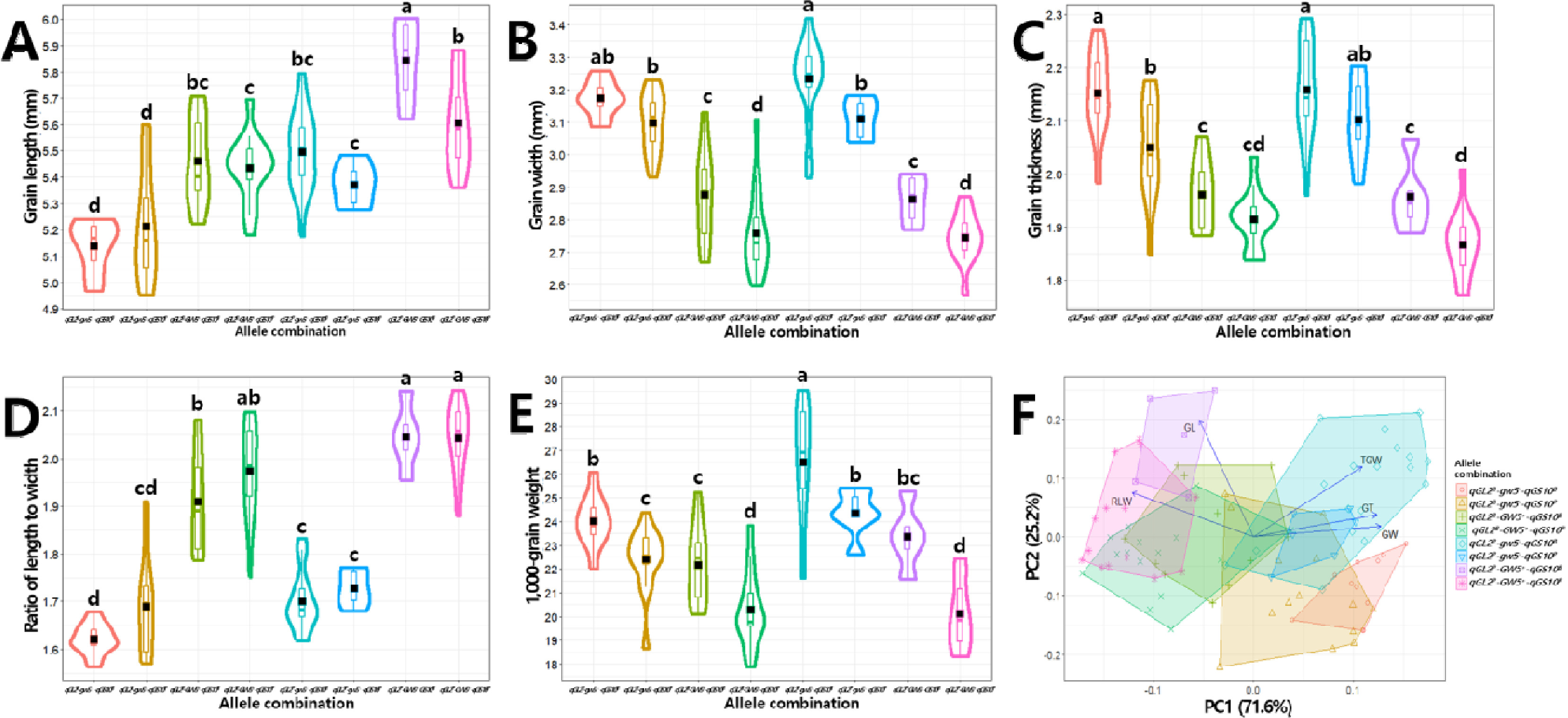
Table 6). ‘보람찬’과 같은 대립유전자형 조합인
qGL2B-gw5-- qGS10B은 현미 길이와 장폭비가 가장 작았으며 너비와 두께는 두번째로 넓고 두꺼웠으며 천립중이 세번째로 무거웠다. ‘보람찬’의 대립유전자형 조합에서
qGL2B가
qGL2P로 치환된 조합인
qGL2P-gw5--qGS10B는 현미 길이가 세번째로 길었고 너비와 두께, 천립중이 8개 조합 중 가장 큰 값을 나타냈다. ‘Pecos’와 같은 대립유전자형 조합인
qGL2P-GW5+-qGS10P는 현미 길이와 장폭비가 두 번째로 크고 너비, 두께, 천립중이 가장 낮은 값을 나타냈으며,
qGS10P가
qGS10B로 치환된 조합인
qGL2P-GW5+-qGS10B는 ‘Pecos’ 대립유전자형 조합에 비해 현미 길이, 너비, 두께, 장폭비 및 천립중이 모두 증가하였다. 대립유전자형 조합들 간 입형 관련 형질 값을 비교 분석하였고, 주성분분석을 통해 입형 관련 형질과 대립유전자형 조합의 구조적 관계를 파악하였다(
Fig. 10). 주성분분석을 통해 전체 입형 관련 표현형 변이의 96.8%를 설명할 수 있었다(
Fig. 10F). 주성분1(표현형 변이의 71.6% 설명)을 기준으로 현미 길이와 장폭비, 두께와 너비 및 천립중이 다른 방향성을 나타냈으며, 대립유전자형 조합의 분포를 살펴본 결과
GW5의 대립유전자형에 따라
GW5+ (Pecos 대립유전자형)를 보유한 4조합(
qGL2P-GW5+-qGS10B,
qGL2P-GW5+-qGS10P,
qGL2B-GW5+-qGS10B,
qGL2B-GW5+- qGS10P)이 현미 길이와 장폭비와 같은 방향에 위치하였으며
gw5- (보람찬 대립유전자형)를 보유한 4조합(
qGL2B-gw5--qGS10B,
qGL2B-gw5--qGS10P,
qGL2P-gw5--qGS10B,
qGL2P-gw5--qGS10P)은 두께와 너비 및 천립중과 같은 방향에 위치하였다. 주성분2(표현형 변이의 25.2%)를 포함하여 대립유전자형 조합들의 구조적 분포를 살펴본 결과,
qGL2P-GW5+-qGS10B와
qGL2P-GW5+-qGS10P 대립유전자형 조합이
qGL2B-gw5--qGS10B와
qGL2B-gw5--qGS10P 조합과 떨어져 분포하고 있어 형질 특성이 다른 것으로 나타났다. 이들 대립유전자형 조합의 분포를 통해 주성분1을 기준으로는
GW5, 주성분2에 대해서는
qGL2 대립유전자형의 영향이 큰 것으로 판단된다.
입형 관련 대립유전자형 조합이 수량 관련 형질에 미치는 영향을 분석하였다. ANOVA 분석 결과,
qGL2는 출수기, 간장, 수장, 수수 및 수당립수,
GW5는 천립중, 등숙률, 제현율, 1주 수량에 가장 큰 영향을 미쳐 이 두 유전자가 수량 관련 형질 변이에 주로 영향을 미쳤으며,
qGS10은 수당립수, 천립중, 등숙률, 1주 수량에 두 번째로 영향을 미쳤다(
Supplementary Table 6). 8개의 대립유전자형 조합 중 모부본인 ‘보람찬’이 보유한
qGL2B-gw5--qGS10B은 천립중과 1주 수량은 높은 편이었으나 등숙률은 72.4%로 낮은 값을 나타냈고, ‘Pecos’의 대립유전자형 조합인
qGL2P-GW5+-qGS10P는 천립중과 1주 수량이 두 번째로 낮은 값을 나타냈으나 등숙률은 80.5%로 중간 순위의 값을 나타냈다(
Table 7). 1주 수량이 가장 높은 조합은 ‘보람찬’이 보유한
gw5을 유지하면서 ‘Pecos’의 대립유전자
qGL2P와
qGS10P가 도입된
qGL2P-gw5--qGS10P로 수량성 뿐만 아니라 등숙률, 제현율, 출수기, 간장, 수당립수가 가장 높은 값을 나타냈다. 그 다음으로 수량성이 높은 조합은
qGL2P-gw5--qGS10B로 천립중은 첫 번째, 1주 수량과 제현율은 두 번째, 등숙률은 세 번째로 높은 특성을 나타냈다. BP_RIL 중 전반적으로 수량성이 높은 조합은 국내 환경조건에서 적응성이 높은 것으로 판단되는
gw5-를 보유하고 있으면서 미국품종인 ‘Pecos’에서 유래한 대립유전자형이 도입된 경우였다. 이를 확인하기 위해 ‘Pecos’ 유래 입형 관련 대립유전자형이 국내 자포니카 벼 품종에 도입되었을 경우에 입형과 수량 관련 형질에 미치는 영향을 분석하였다(
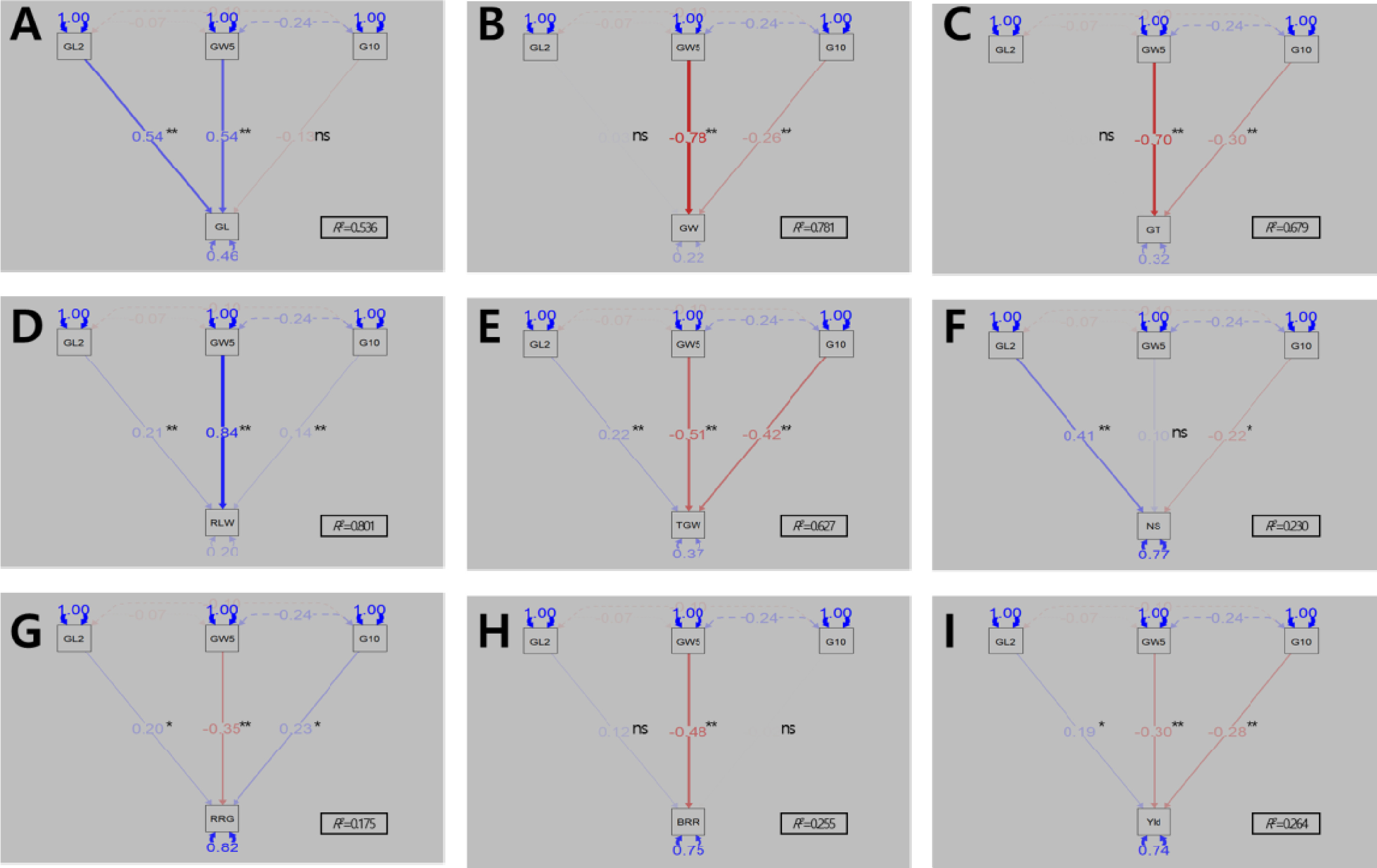
Fig. 11). 경로계수분석 결과, 입형 관련 대립유전자형들이 입형 관련 형질 변이를 설명하는 정도를 나타내는 결정계수(
R2)는 0.536-0.801이었다. 현미 길이에 대해서는
qGL2P와
GW5+가 비슷한 수준(표준화 회귀계수 0.54)으로 증가하는 방향으로 작용하였고, 너비와 두께에는
GW5+ (-0.78, -0.70)와
qGS10P (-0.26, -0.30)가 감소하는 방향, 장폭비에는
GW5+ (0.84),
qGL2P (0.21),
qGS10P (0.14) 모두 증가하는 방향으로 작용하였으며, 천립중에는
GW5+ (-0.51)와
qGS10P (-0.42)는 감소,
qGL2P (0.22)는 증가하는 방향으로 작용하였다. 이를 통해 ‘보람찬’과 같은 입형 관련 대립유전자형 조합을 가진 국내품종에 미국품종 ‘Pecos’의 대립유전자형이 도입될 시 현미 길이와 장폭비는 증가하고 너비와 두께 및 천립중은 전반적으로 감소하는 경향이나,
qGL2P 도입의 경우 천립중이 증가하는 방향으로도 작용할 수 있음을 확인하였다. 수량 관련 형질에 대한 결정계수(
R2)는 0.175-0.264로 입형 관련 대립유전자형들로 설명되는 정도가 낮았으나 도입 시 형질 변이를 살펴보면, 수당립수에 대해서는
qGL2P는 증가(표준화 회귀계수 0.41),
qGS10P (-0.22)는 감소에 영향을 미쳤고, 등숙률에는
qGL2P (0.20)와
qGS10P (0.23)는 증가,
GW5+ (-0.35)는 감소하는 방향, 제현율은
GW5+ (-0.48)가 감소, 1주 수량에는
GW5+ (-0.30)와
qGS10P (-0.28)는 감소,
qGL2P (0.19)는 증가하는 방향으로 작용하였다. 이를 통해 미국품종 ‘Pecos’가 보유하고 있는
GW5+의 도입은 등숙률, 제현율, 1주 수량이 감소하여 수량성에 부정적인 영향을 미칠 수 있고,
qGS10P의 도입은 등숙률은 증가하나 수당립수와 1주 수량이 감소하게 되어 수량성 향상에 제한적일 수 있으며,
qGL2P의 도입은 수당립수가 증가함에도 등숙률이 향상되고 1주 수량도 증가하여 수량성에 긍정적인 영향을 미칠 것으로 판단된다.
우리나라 자포니카 벼 품종들은 온대 자포니카 벼에 속하며 대부분 협소한 유전적 배경을 가지고 있는 것으로 알려져 있다(
Lee et al. 2022). 반면 미국 재배품종들은 주로 열대 자포니카를 이용하여 육종되었기 때문에 온대 자포니카 벼와 유전 조성이 다른 품종들이 많은 것으로 보고되었다(
Lu et al. 2005). 우리나라 자포니카 벼 품종의 수량 및 입형 관련 형질에 대한 유전적 다양성을 확대하기 위하여 국내 온대 자포니카 중만생 초다수 품종 ‘보람찬’과 열대 자포니카이면서 중립종 입형 특성을 가진 조생종 미국품종 ‘Pecos’를 교배하여 재조합 자식 집단을 작성하고 수량 및 입형 관련 형질에 대한 QTL 분석을 수행하였다. 이를 통해 수량과 입형 관련 QTL과 후보 유전자를 탐색하였으며 이들이 형질 변이에 미치는 영향을 분석하였다. 탐색된 출수기 관련 주동 유전자
Hd1은 출수기 뿐만 아니라 간장, 수장, 수당립수, 천립중, 제현율, 등숙률, 1주 수량 등 수수를 제외한 모든 수량구성요소에 영향을 미치는 다면발현 현상을 나타냈다. 이와 함께 출수기 관련 미동 유전자
Hd6가 탐색되었고, 두 개의 출수기 관련 유전자
Hd1과
Hd6의 상호작용을 통해 출수일수를 30일 이상 조절할 수 있었으며, 대립유전자형 조합이 수당립수와 1주 수량 등 수량구성요소 변이에 미치는 영향을 분석하였다. 입형 관련해서는
qGL2,
GW5,
qGS10 세 개의 QTL 및 유전자를 탐색하였다. 우리나라 자포니카 벼 품종들이 대부분 가지고 있는 것으로 보고된
gw5- 대립유전자를 보유한 조합이 수량성 측면에서 전반적으로 양호한 특성을 나타내 국내 환경 조건에서 적응성이 높은 것으로 판단되었다. 여기에 ‘Pecos’가 보유한 입형 관련 대립유전자
qGL2P와
qGS10P의 도입은 입형을 다양화하고 수당립수, 등숙률, 1주 수량 향상에 긍정적인 영향을 미치는 것으로 확인되어,
qGL2P-gw5--qGS10B,
qGL2P-gw5--qGS10P와 같은 새로운 대립유전자형 조합은 우리나라 자포니카 벼 품종의 수량과 입형 관련 형질의 유전적 개선에 도움이 될 것으로 기대된다. 본 연구를 통해 파악된 유전 양상과 관련 유전자들의 작용은 육종사업에 반영하여 다양한 입형 특성을 가지면서 수량성이 향상된 자포니카 벼 품종을 효과적으로 개발하는데 활용할 계획이다.
적요
‘보람찬’은 우리나라 온대 자포니카 중만생종 초다수 벼 품종이고 ‘Pecos’는 열대 자포니카이면서 중립종 입형 특성을 가진 조생종 미국품종이다. 우리나라 자포니카 벼 품종의 수량 및 입형 관련 형질에 대한 유전적 다양성을 확대하기 위하여 ‘보람찬’과 ‘Pecos’를 이용한 재조합 자식 집단을 육성하고 수량 및 입형 관련 형질에 대한 QTL 분석을 수행하였다. 수량 관련 형질 중 출수기에 대한 QTL이 3번과 6번 염색체에서 탐색되었고 후보 유전자로 Hd6와 Hd1이 추정되었다. 탐색된 출수기 관련 주동 유전자 Hd1은 출수기 뿐만 아니라 간장, 수장, 수당립수, 천립중, 제현율, 등숙률, 1주 수량 등 대부분의 수량구성요소에 영향을 미치는 다면발현(pleiotropy) 현상을 나타냈다. 출수기 관련 미동 유전자 Hd6는 주동 유전자 Hd1의 대립유전자형에 따라 출수기와 수당립수에 미치는 영향이 가려지는 masking 현상이 나타났다. Hd1과 Hd6의 대립유전자형 조합에 따라 출수일수를 30일 가량 조절할 수 있었으며, 출수가 지연되는 정상기능형 대립유전자가 집적될수록 수량 관련 형질 값이 증가하였다. 입형 관련하여 2번, 5번, 10번 염색체에서 qGL2, GW5, qGS10 세 개의 QTL 및 유전자가 탐색되었다. GW5는 현미 길이, 너비, 두께, 장폭비 및 천립중 등 입형 관련 모든 형질에 관여하였으며, 표현형 변이에 미치는 영향이 큰 주동 유전자로 확인되었다. qGL2는 현미 길이에 관여하는 QTL로 수당립수 관련 qNS2와 동일위치에서 탐색되었다. qGS10는 현미 너비, 두께, 천립중에 관여하는 QTL로 확인되었다. 입형 관련 대립유전자형 조합은 입형 뿐만 아니라 수량 관련 형질 변이에도 영향을 미쳤다. ‘보람찬’의 gw5- 대립유전자를 보유한 조합이 수량성 측면에서 전반적으로 양호한 특성을 나타냈다. 여기에 ‘Pecos’의 qGL2P, qGS10P 대립유전자가 도입될 경우 입형을 다양화하고 수당립수, 등숙률, 1주 수량 향상에 긍정적인 영향을 미치는 것으로 확인되었다. qGL2P-gw5--qGS10B, qGL2P-gw5--qGS10P와 같은 새로운 대립유전자형 조합은 우리나라 자포니카 벼 품종의 수량 및 입형 관련 유전적 다양성을 개선하는데 기여할 것으로 기대된다.
보충자료
본문의 Supplementary Tables 1-6 및 Supplementary Fig. 1은 한국육종학회지 홈페이지에서 확인할 수 있습니다.
사사
본 논문은 농촌진흥청 작물시험연구사업(남부지역 적응 밥쌀용 재배안정성 벼 품종개발(3단계), 과제번호: PJ0160670 62024)의 지원으로 수행된 결과의 일부입니다.
Fig. 1Phenotypic distribution of yield-related traits in the population of 94 RILs derived from a cross between ‘Boramchan’ and ‘Pecos’ across two years (A-I). The blue and yellow curves and dashed lines represent the density plots and mean values of BP_RIL for the years 2022 and 2023, respectively. The blue and yellow inverted triangles represent the mean values of parents for the years 2022 and 2023, respectively. BRC: Boramchan, PCS: Pecos.
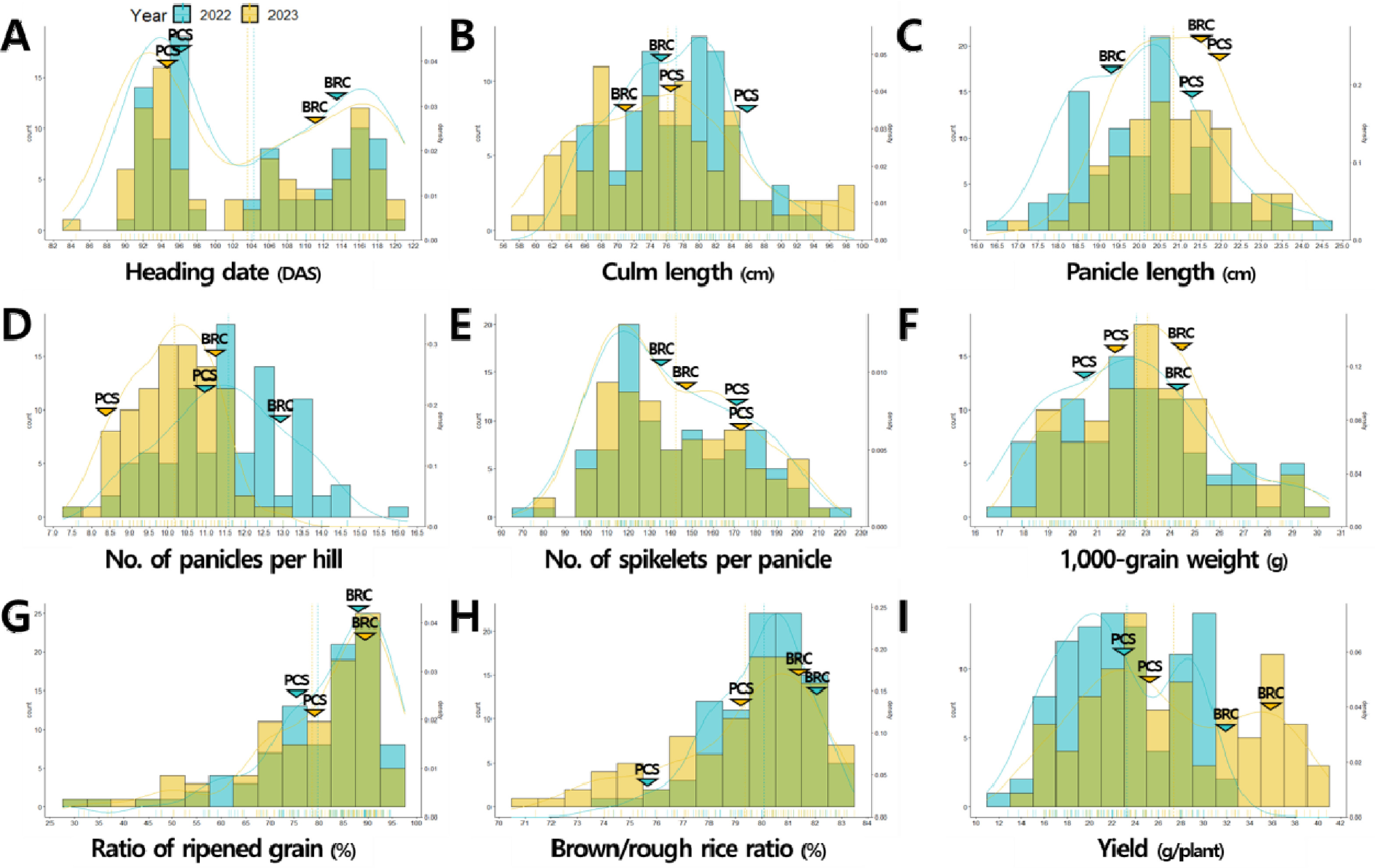
Fig. 2Phenotypic distribution of grain-related traits in the population of 94 RILs derived from a cross between ‘Boramchan’ and ‘Pecos’ across two years (A-D) and path analysis of the causal relationship among grain-related traits to explain 1,000-grain weight variation in 2022 (E) and 2023 (F). The blue and yellow curves and dashed lines represent the density plots and mean values of BP_RIL for the years 2022 and 2023, respectively. The blue and yellow inverted triangles represent the mean values of parents for the years 2022 and 2023, respectively. BRC: Boramchan, PCS: Pecos. GL: grain length, GW: grain width, GT: grain thickness, TGW: 1,000-grain weight. Single-headed continuous arrows represent linear dependencies (direct effects) calculated as standardized regression coefficients, while double-headed dotted arrows indicate correlation coefficients between each traits. Blue and red arrows indicate positive and negative effects, respectively. The arrow line thickness represents the proportion of the effect. ** indicates significant at 1% probability level.
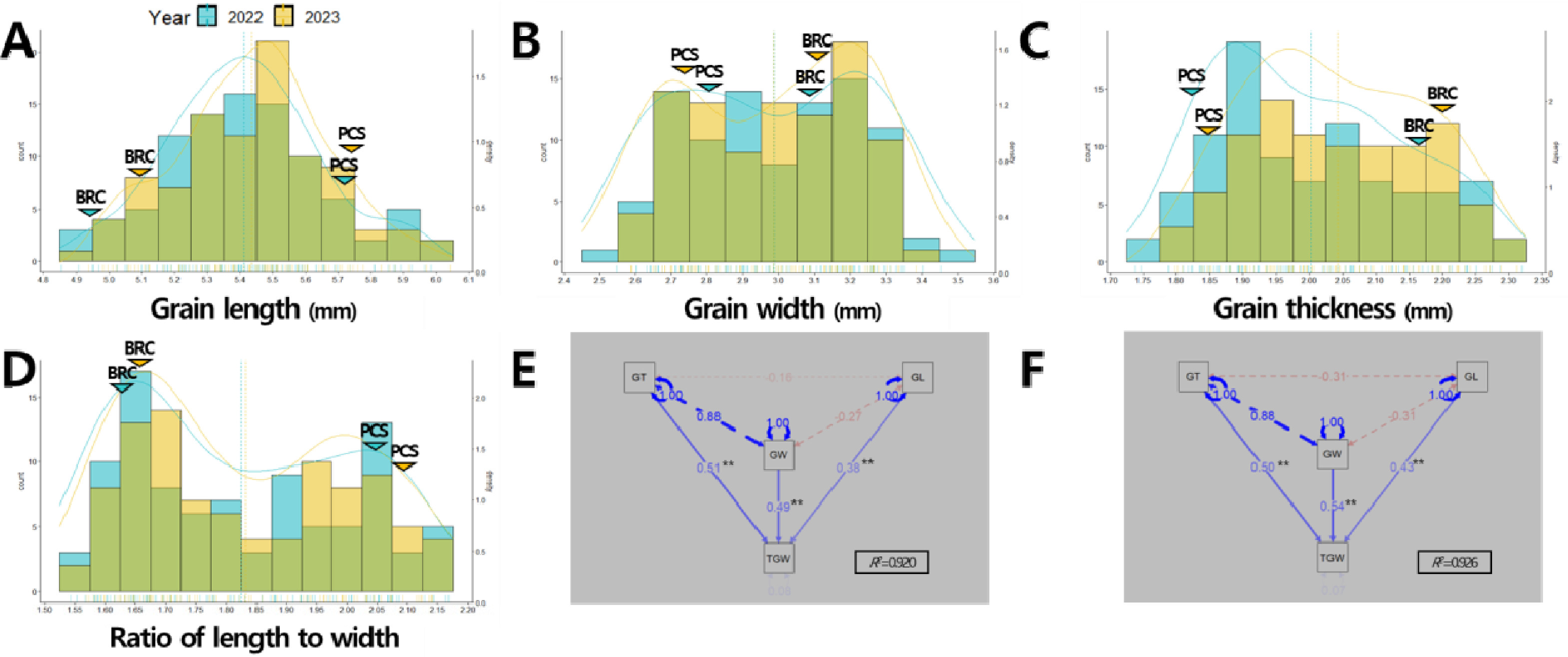
Fig. 3Relationship among yield-related traits. Correlation analysis in 2022 (A) and 2023 (B). Path analysis of the causal relationship among yield-related traits to explain yield variation in 2022 (C) and 2023 (D). HD: heading date, CL: culm length, PL: panicle length, PN: number of panicles per hill, NS: number of spikelets per panicle, TGW: 1,000-grain weight, RRG: ratio of ripened grain, BRR: brown/rough rice ratio, Yld: yield. Single-headed continuous arrows represent linear dependencies (direct effects) calculated as standardized regression coefficients, while double-headed dotted arrows indicate correlation coefficients between each traits. Blue and red arrows indicate positive and negative effects, respectively. The arrow line thickness represents the proportion of the effect. Ns and ** indicate no significant and significant at 1% probability level, respectively.
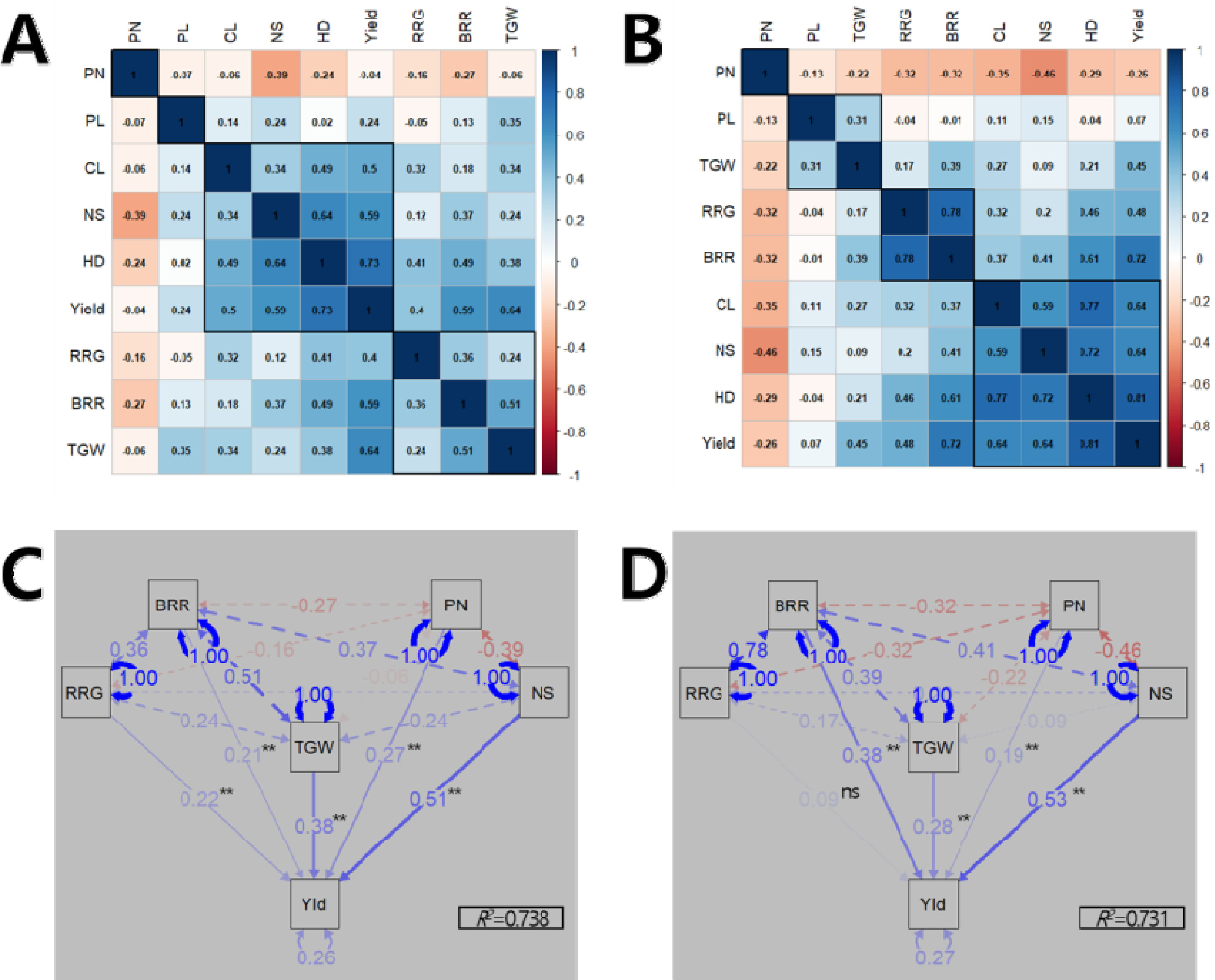
Fig. 4Genetic map showing the LOD scores and locations of QTLs associated with yield-related traits in the population of 94 RILs derived from a cross between ‘Boramchan’ and ‘Pecos’ across two years. A: heading date (HD), B: culm length (CL), C: number of spikelets per panicle (NS), D: brown/rough rice ratio (BRR), E: yield. The red and green curves represent LOD scores for the year 2022 and 2023, respectively. The red and green horizontal lines and symbols indicate the positions and names of QTLs for the years 2022 and 2023, respectively.
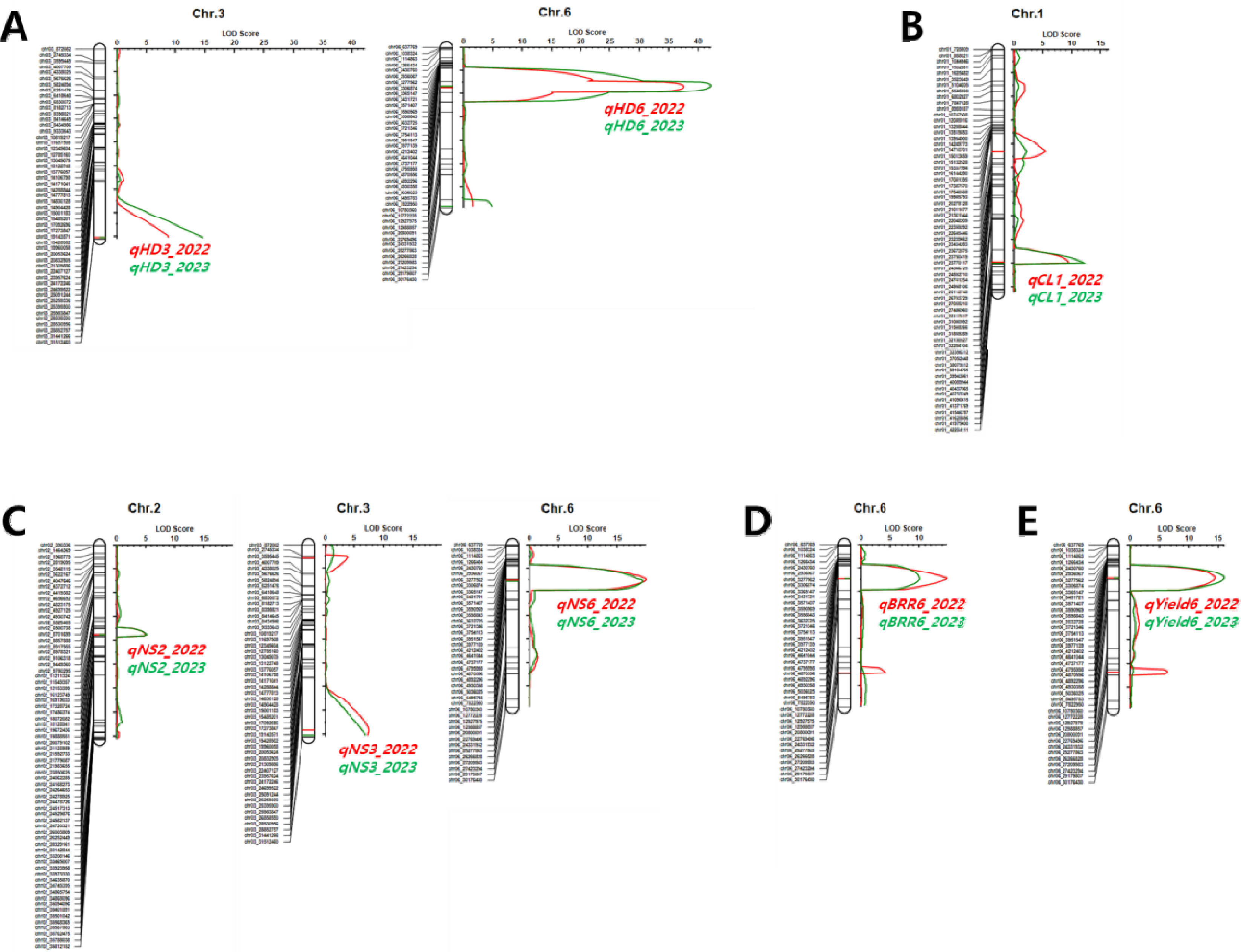
Fig. 5Genetic map showing the LOD scores and locations of QTLs associated with grain-related traits in the population of 94 RILs derived from a cross between ‘Boramchan’ and ‘Pecos’ across two years. A: grain length (GL), B: grain width (GW), GT: grain thickness (GT), D: ratio of length to width (RLW), E: 1,000-grain weight (TGW). The red and green curves represent LOD scores for the year 2022 and 2023, respectively. The red and green horizontal lines and symbols indicate the positions and names of QTLs for the years 2022 and 2023, respectively.
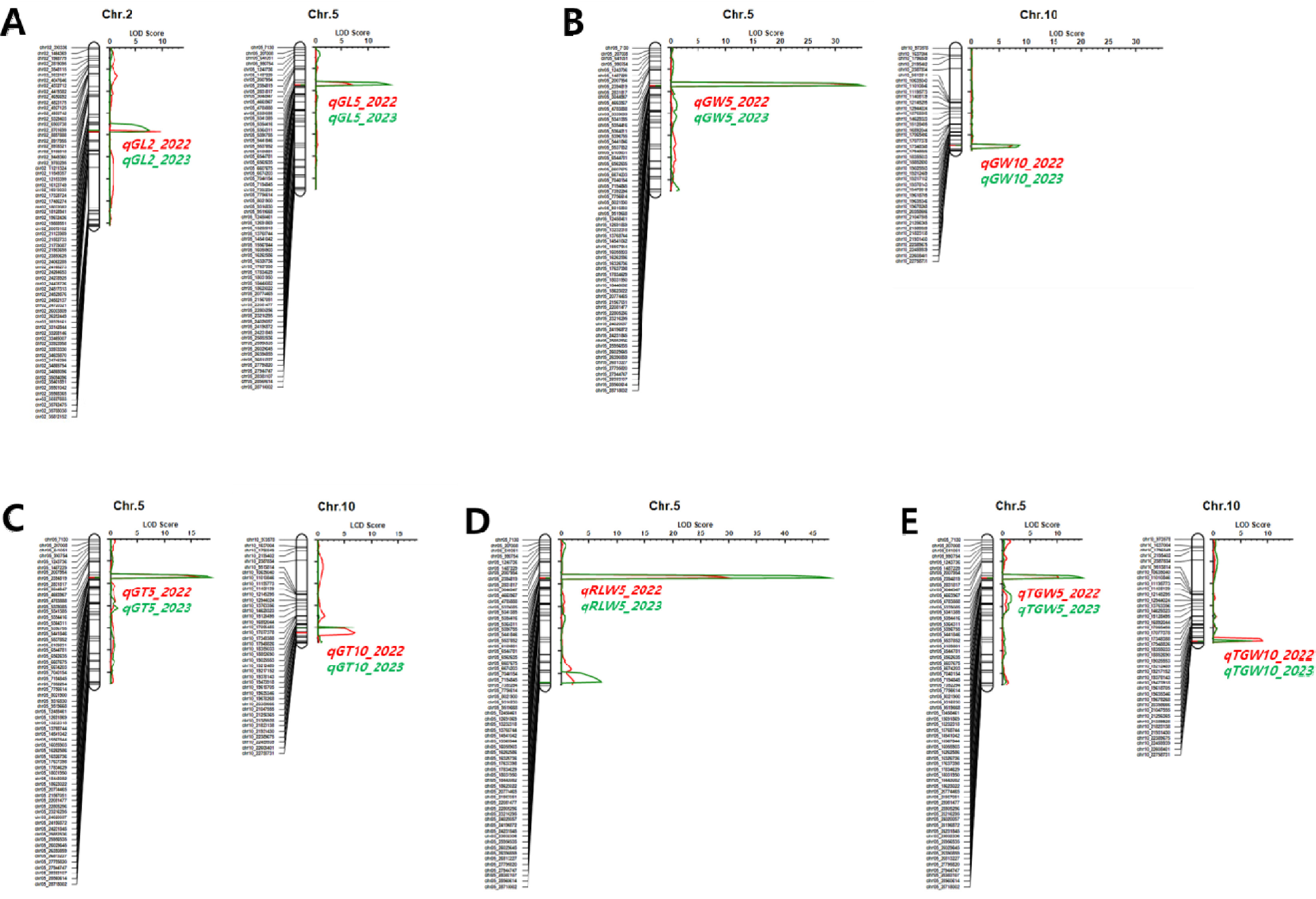
Fig. 6Effects of Hd1 alleles on the phenotypes of yield-related traits in the BP_RIL population. Violin plots showing the variation in heading date (A), culm length (B), panicle length (C), number of panicles per hill (D), number of spikelets per panicle (E), 1,000-grain weight (F), ratio of ripened grain (G), brown/rough rice ratio (H), and yield (I) by allele types, Hd1+ (Boramchan type) and hd1- (Pecos type). The black rectangles indicate the means of traits. NS, **, and, *** indicate no significant, significance at the 0.01 and 0.001 probability levels by t-test, respectively.
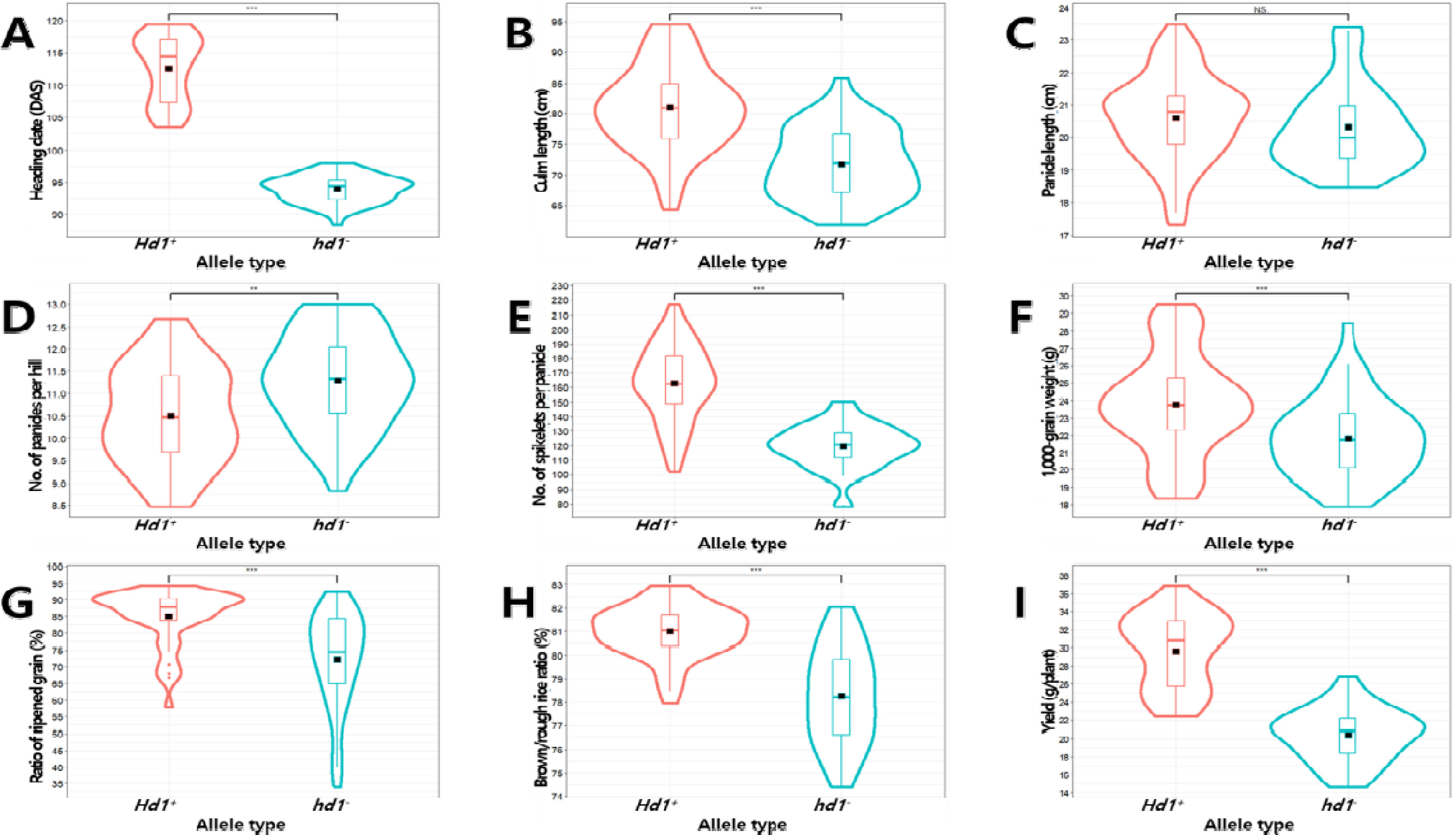
Fig. 7Effects of Hd1 and Hd6 allele combinations on the phenotype of yield-related traits in the BP_RIL population. Violin plots showing the variation in heading date (A), culm length (B), panicle length (C), number of panicles per hill (D), number of spikelets per panicle (E), 1,000-grain weight (F), ratio of ripened grain (G), brown/rough rice ratio (H), and yield (I) by allele combinations. The black rectangles indicate the means of traits. The same letters on the violin plots indicate that the not significantly different at p<0.05 (ANOVA followed by DMRT).
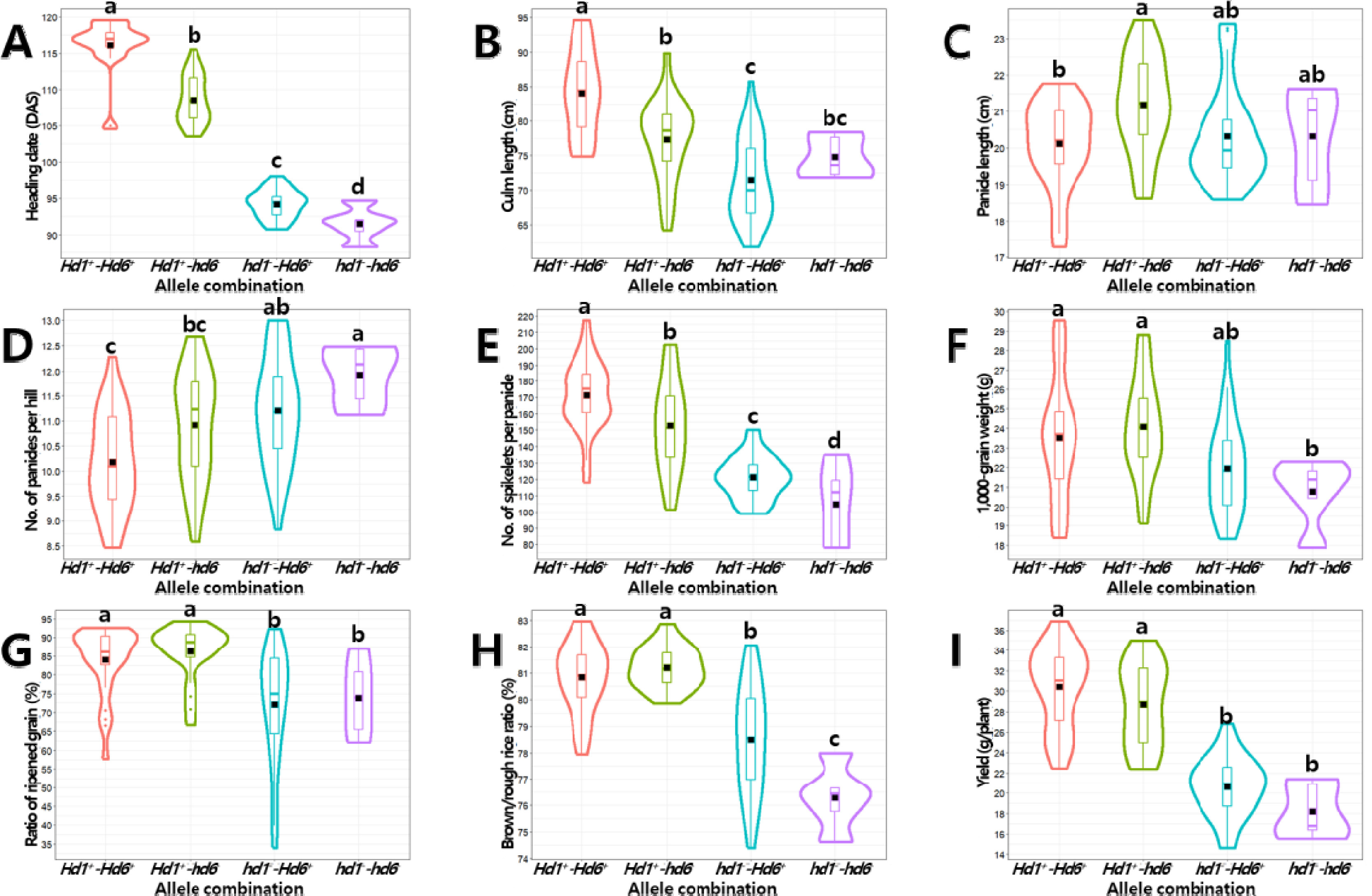
Fig. 8Interaction effects of Hd1 and Hd6 alleles on yield-related traits (A-I). A: heading date, B: culm length, C: panicle length, D: number of panicles per hill, E: number of spikelets per panicle, F: 1,000-grain weight, G: ratio of ripened grain, H: brown/rough rice ratio, I: yield.
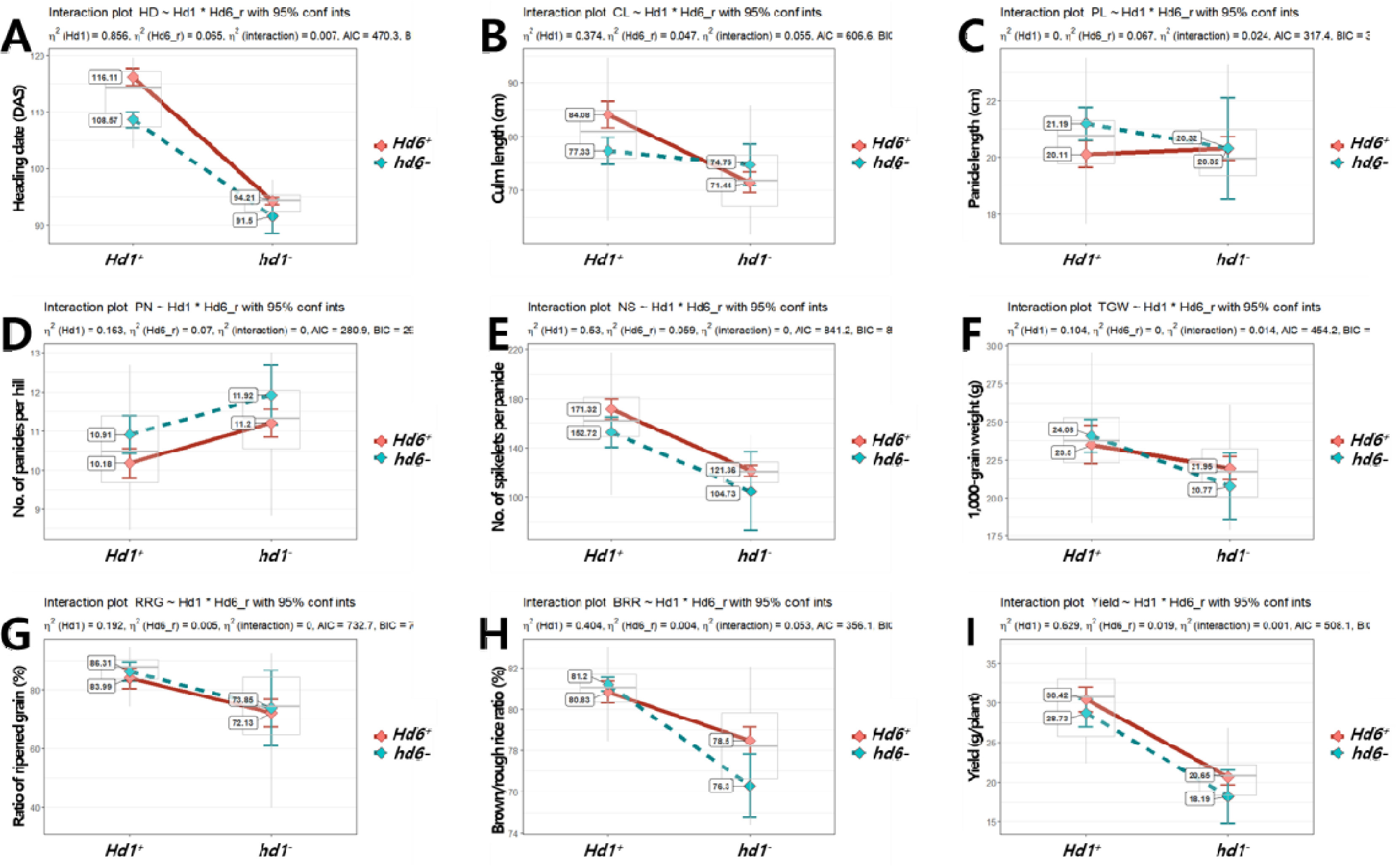
Fig. 9Effects of qGL2 (A), GW5 (B), and qGS10 (C) alleles on the phenotype of grain-related traits in the BP_RIL population. Violin plots showing the variation in grain length, grain width, grain thickness, ratio of length to width, and 1,000-grain weight by allele types of ‘Boramchan’ (qGL2B, gw5-, qGS10B) and ‘Pecos’ (qGL2P, GW5+, qGS10P). The black rectangles indicate the means of traits. NS, **, and, *** indicate no significant, significance at the 0.01 and 0.001 probability levels by t-test, respectively.
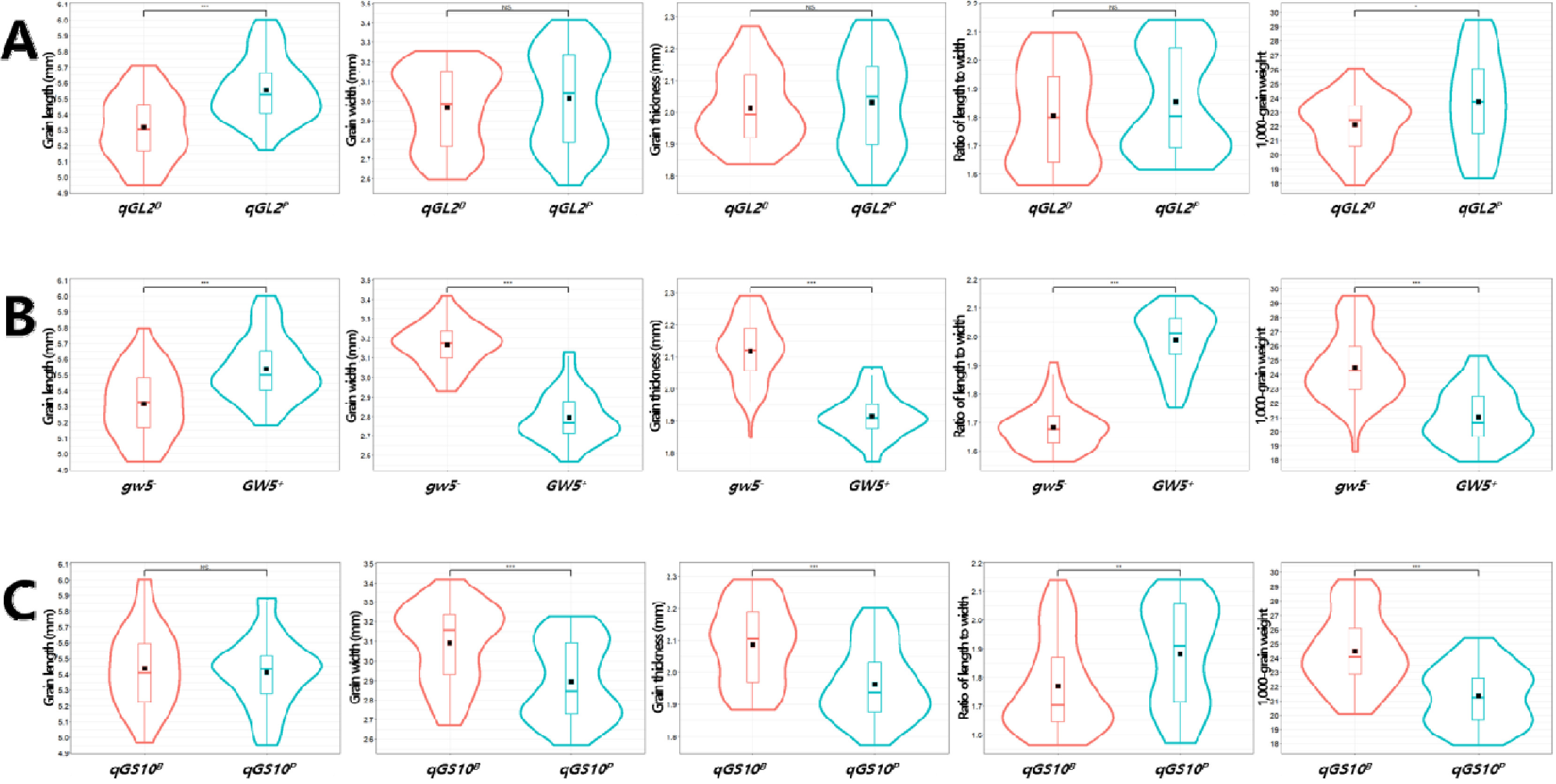
Fig. 10Effects of qGL2, GW5, and qGS10 allele combinations on the phenotype of grain-related traits in the BP_RIL population (A-E) and principal component analysis (PCA, F). Violin plots showing the variation in grain length (GL, A), grain width (GW, B), grain thickness (GT, C), ratio of length to width (RLW, D), 1,000-grain weight (TGW, F) by allele combinations. The black rectangles indicate the means of traits. The same letters on the violin plots indicate that the not significantly different at p<0.05 (ANOVA followed by DMRT). In PCA, BP_RILs were classified by allele combinaitons. PC1: principal component 1, PC2: principal component 2.
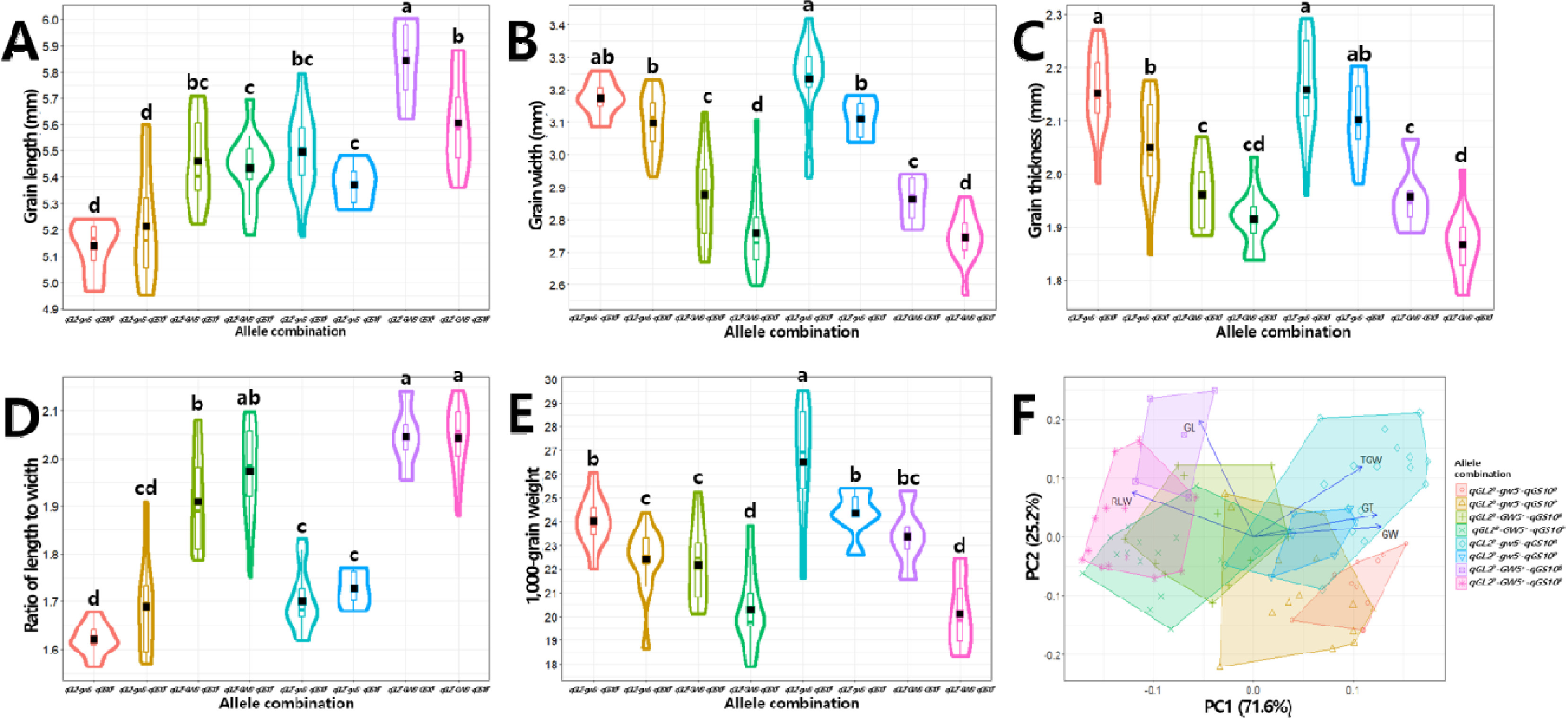
Fig. 11Introgression effect of qGL2 (GL2), GW5, and qGS10 (G10) alleles of ‘Pecos’ on grain and yield-related traits. A: grain length (GL), B: grain width (GW), C: grain thickness (GT), D: ratio of length to width (RLW), E: 1,000-grain weight (TGW), F: number of spikelet per panicle (NS), G: ratio of ripened grain (RRG), H: brown/rough rice ratio (BRR), I: yield (Yld). Single-headed continuous arrows represent linear dependencies (direct effects) calculated as standardized regression coefficients, while double-headed dotted arrows indicate correlation coefficients between each trait. Blue and red arrows indicate positive and negative effects, respectively. The arrow line thickness represents the proportion of the effect. Ns, *, and ** indicate no significant and significant at 5% and 1% probability level, respectively.
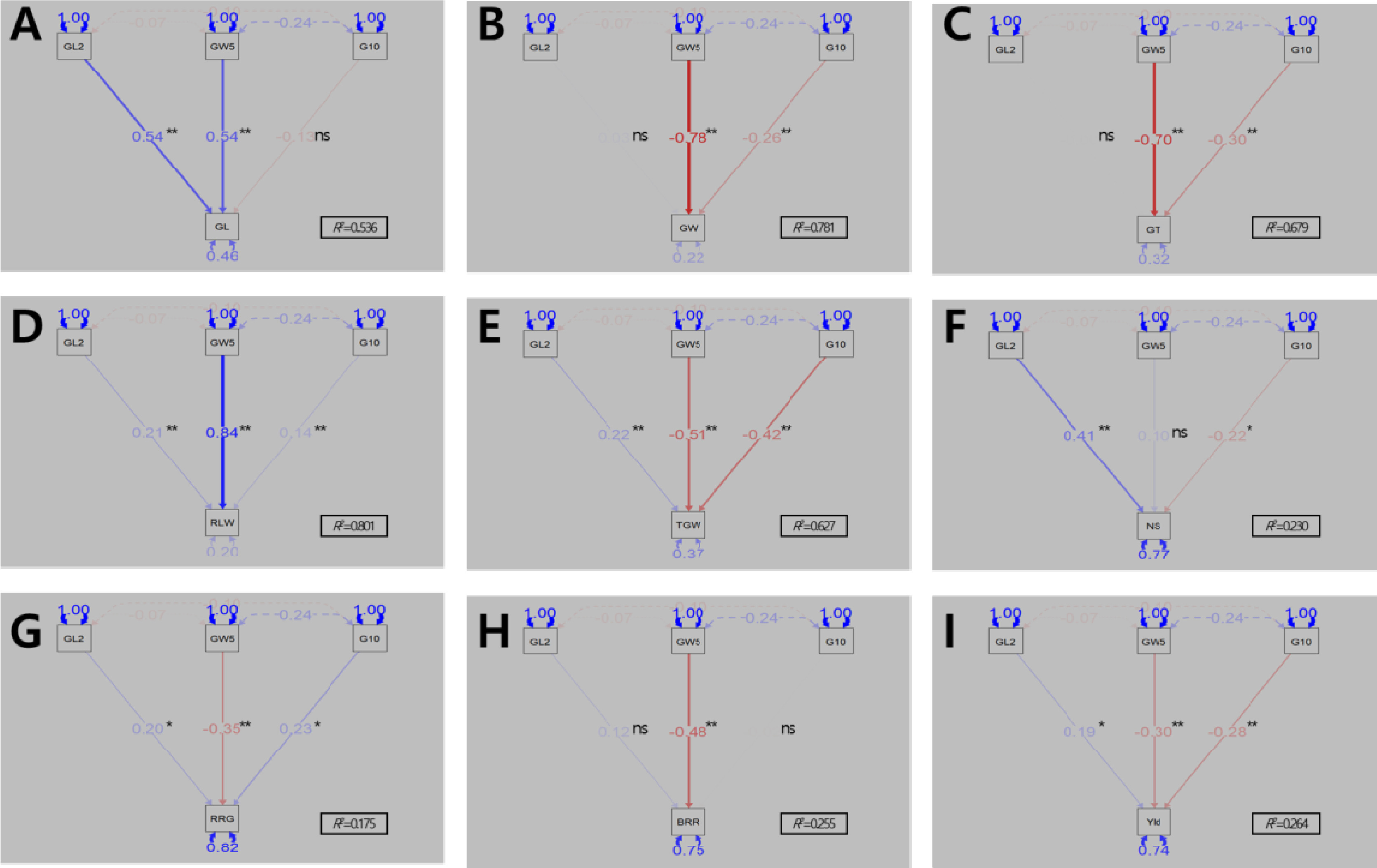
Table 1QTLs associated with yield-related traits identified in the recombinant inbred line population derived from a cross between ‘Boramchan’ (P1) and ‘Pecos’ (P2) during the years 2022-2023.
Table 1
|
Traitz
|
QTL
name |
Chr.y
|
Position
(cM) |
Interval-flanking markers |
LODx
|
PVEw
(%) |
Addv
|
Candidate
geneu
|
|
|
Left |
Right |
|
HD |
qHD3_2022
|
3 |
161 |
chr03_28852757 |
chr03_31441266 |
8.7 |
8.8 |
-3.54 |
Hd6
|
|
qHD6_2022
|
6 |
34 |
chr06_7822950 |
chr06_10780360 |
37.6 |
65.4 |
9.13 |
Hd1
|
|
qHD3_2023
|
3 |
161 |
chr03_28852757 |
chr03_31441266 |
14.7 |
8.4 |
-3.99 |
Hd6
|
|
qHD6_2026
|
6 |
33 |
chr06_7822950 |
chr06_10780360 |
42.2 |
72.9 |
10.83 |
Hd1
|
|
CL |
qCL1_2022
|
1 |
179 |
chr01_37052448 |
chr01_38079312 |
9.4 |
22.2 |
-3.21 |
SD1
|
|
qCL1_2023
|
1 |
180 |
chr1_37052448 |
chr1_38079312 |
12.0 |
20.6 |
-4.83 |
SD1
|
|
NS |
qNS2_2022
|
2 |
75 |
chr02_18128941 |
chr02_19672436 |
5.0 |
6.7 |
-9.55 |
- |
|
qNS3_2022
|
3 |
156 |
chr03_28852757 |
chr03_31441266 |
7.3 |
16.0 |
-16.08 |
Hd6
|
|
qNS6_2022
|
6 |
30 |
chr06_7822950 |
chr06_10780360 |
19.9 |
40.4 |
23.42 |
Hd1
|
|
qNS2_2023
|
2 |
75 |
chr02_18128941 |
chr02_19672436 |
5.1 |
8.1 |
-9.57 |
- |
|
qNS3_2023
|
3 |
161 |
chr03_28852757 |
chr03_31441266 |
6.8 |
11.9 |
-12.61 |
Hd6
|
|
qNS6_2023
|
6 |
31 |
chr06_7822950 |
chr06_10780360 |
19.3 |
44.8 |
22.41 |
Hd1
|
|
BRR |
qBRR6_2022
|
6 |
29 |
chr06_5495783 |
chr06_7822950 |
14.7 |
23.0 |
1.14 |
Hd1
|
|
qBRR6_2023
|
6 |
29 |
chr06_5495783 |
chr06_7822950 |
10.1 |
25.0 |
1.25 |
Hd1
|
|
Yield |
qYield6_2022
|
6 |
28 |
chr06_5495783 |
chr06_7822950 |
14.3 |
42.9 |
2.82 |
Hd1
|
|
qYield6_2023
|
6 |
28 |
chr06_5495783 |
chr06_7822950 |
16.0 |
40.8 |
3.79 |
Hd1
|
Table 2QTLs associated with grain-related traits identified in the recombinant inbred line population derived from a cross between ‘Boramchan’ (P1) and ‘Pecos’ (P2) during the years 2022-2023.
Table 2
|
Traitz
|
QTL
name |
Chr.y
|
Position
(cM) |
Interval-flanking markers |
LODx
|
PVEw
(%) |
Addv
|
Candidate
geneu
|
|
|
Left |
Right |
|
GL |
qGL2_2022
|
2 |
76 |
chr02_19672436 |
chr02_19888551 |
8.7 |
27.0 |
-0.12 |
OML4 (LARGE1/EHD6)
|
|
qGL5_2022
|
5 |
34 |
chr05_4783888 |
chr05_5339085 |
6.8 |
21.1 |
-0.10 |
qSW5/GW5
|
|
qGL2_2023
|
2 |
75 |
chr02_18128941 |
chr02_19672436 |
7.4 |
13.5 |
-0.08 |
OML4 (LARGE1/EHD6)
|
|
qGL5_2023
|
5 |
34 |
chr05_4783888 |
chr05_5339085 |
14.0 |
31.3 |
-0.13 |
qSW5/GW5
|
|
GW |
qGW5_2022
|
5 |
34 |
chr05_4783888 |
chr05_5339085 |
31.2 |
68.5 |
0.18 |
qSW5/GW5
|
|
qGW10_2022
|
10 |
89 |
chr10_21256365 |
chr10_21389928 |
7.5 |
6.9 |
0.06 |
GW10 etc.
|
|
qGW5_2023
|
5 |
34 |
chr05_4783888 |
chr05_5339085 |
34.8 |
69.9 |
0.16 |
qSW5/GW5
|
|
qGW10_2023
|
10 |
89 |
chr10_21256365 |
chr10_21389928 |
8.8 |
8.2 |
0.05 |
GW10 etc.
|
|
GT |
qGT5_2022
|
5 |
34 |
chr05_4783888 |
chr05_5339085 |
16.5 |
47.5 |
0.09 |
qSW5/GW5
|
|
qGT10_2022
|
10 |
85 |
chr10_20358666 |
chr10_21047555 |
6.8 |
15.0 |
0.05 |
GW10 etc.
|
|
qGT5_2023
|
5 |
34 |
chr05_4783888 |
chr05_5339085 |
18.5 |
48.9 |
0.08 |
qSW5/GW5
|
|
qGT10_2023
|
10 |
80 |
chr10_19473918 |
chr10_19618705 |
5.8 |
11.5 |
0.04 |
GW10 etc.
|
|
RLW |
qRLW5_2022
|
5 |
34 |
chr05_4783888 |
chr05_5339085 |
29.9 |
71.6 |
-0.14 |
qSW5/GW5
|
|
qRLW5_2023
|
5 |
34 |
chr05_4783888 |
chr05_5339085 |
47.8 |
76.2 |
-0.16 |
qSW5/GW5
|
|
TGW |
qTGW5_2022
|
5 |
34 |
chr05_4783888 |
chr05_5339085 |
10.3 |
26.3 |
1.24 |
qSW5/GW5
|
|
qTGW10_2022
|
10 |
92 |
chr10_21823138 |
chr10_21931430 |
9.0 |
21.6 |
1.13 |
GW10 etc.
|
|
qTGW5_2023
|
5 |
34 |
chr05_4783888 |
chr05_5339085 |
14.6 |
42.9 |
1.64 |
qSW5/GW5
|
|
qTGW10_2023
|
10 |
92 |
chr10_21823138 |
chr10_21931430 |
6.1 |
15.0 |
0.97 |
GW10 etc.
|
Table 3Effects of Hd1, Hd6, and SD1 alleles on the phenotype of yield-related traits.
Table 3
|
Allele typez
|
n |
HDy
(DAS) |
CL
(cm) |
PL
(cm) |
PN |
NS |
TGW
(g) |
RRG
(%) |
BRR
(%) |
Yield
(g/plant) |
|
Hd1+
|
50 |
113**
|
81**
|
20.6ns
|
10.5**
|
163**
|
23.8**
|
85.1**
|
81.0**
|
29.6**
|
|
hd1-
|
44 |
94 |
72 |
20.3 |
11.3 |
119 |
21.8 |
72.3 |
78.2 |
20.4 |
|
hd6-
|
28 |
106ns
|
77ns
|
21.0*
|
11.1ns
|
144ns
|
23.5ns
|
84.1**
|
80.3ns
|
26.8ns
|
|
Hd6+
|
66 |
103 |
77 |
20.2 |
10.8 |
142 |
22.6 |
77.0 |
79.5 |
24.6 |
|
sd1-
|
47 |
103ns
|
73**
|
20.6ns
|
10.9ns
|
143ns
|
23.1ns
|
79.4ns
|
80.1ns
|
25.4ns
|
|
SD1+
|
47 |
105 |
80 |
20.4 |
10.9 |
142 |
22.6 |
78.8 |
79.3 |
25.2 |
Table 4Effects of Hd1 and Hd6 allele combinations on the phenotype of yield-related traits.
Table 4
|
Allele combinationz
|
n |
HDy
(DAS) |
CL
(cm) |
PL
(cm) |
PN |
NS |
TGW
(g) |
RRG
(%) |
BRR
(%) |
Yield
(g/plant) |
|
Hd1+-Hd6+
|
27 |
116ax
|
84a |
20.1b |
10.2c |
171a |
23.5a |
84.0a |
80.8a |
30.4a |
|
Hd1+-hd6-
|
23 |
109b |
77b |
21.2a |
10.9bc |
153b |
24.1a |
86.3a |
81.2a |
28.7a |
|
hd1--Hd6+
|
39 |
94c |
71c |
20.3ab |
11.2ab |
121c |
22.0ab |
72.1b |
78.5b |
20.7b |
|
hd1--hd6-
|
5 |
92d |
75bc |
20.3ab |
11.9a |
105d |
20.8b |
73.9b |
76.3c |
18.2b |
Table 5Effects of qGL2, GW5, and qGS10 alleles on the phenotype of yield-related traits.
Table 5
|
Allele typez
|
n |
HDy
(DAS) |
CL
(cm) |
PL
(cm) |
PN |
NS |
TGW
(g) |
RRG
(%) |
BRR
(%) |
Yield
(g/plant) |
|
qGL2B
|
51 |
101**
|
74**
|
20.1**
|
11.2**
|
130**
|
22.1**
|
76.7*
|
79.4ns
|
24.0*
|
|
qGL2P
|
43 |
107 |
80 |
20.9 |
10.5 |
157 |
23.8 |
82.0 |
80.1 |
26.8 |
|
gw5-
|
49 |
106*
|
77ns
|
20.6ns
|
10.8ns
|
142ns
|
24.5**
|
82.9**
|
80.7**
|
27.4**
|
|
GW5+
|
45 |
101 |
76 |
20.4 |
10.9 |
143 |
21.0 |
74.9 |
78.6 |
23.0 |
|
qGS10B
|
45 |
106*
|
78*
|
20.8*
|
10.8ns
|
150*
|
24.5**
|
77.4ns
|
80.0ns
|
27.6**
|
|
qGS10P
|
49 |
102 |
75 |
20.2 |
10.9 |
136 |
21.4 |
80.6 |
79.4 |
23.2 |
Table 6Effects of qGL2, GW5, and qGS10 allele combinations on the phenotype of grain-related traits.
Table 6
|
Allele combinationz
|
n |
GLy
|
GW |
GT |
RLW |
TGW |
|
qGL2B-gw5--qGS10B
|
11 |
5.14dx
|
3.17ab |
2.15a |
1.62d |
24.0b |
|
qGL2B-gw5--qGS10P
|
14 |
5.21d |
3.10b |
2.05b |
1.69cd |
22.4c |
|
qGL2B-GW5+-qGS10B
|
11 |
5.46bc |
2.88c |
1.96c |
1.91b |
22.2c |
|
qGL2B-GW5+-qGS10P
|
15 |
5.43c |
2.76d |
1.91cd |
1.97ab |
20.3d |
|
qGL2P-gw5--qGS10B
|
18 |
5.50bc |
3.23a |
2.16a |
1.70c |
26.5a |
|
qGL2P-gw5--qGS10P
|
6 |
5.37c |
3.11b |
2.10ab |
1.73c |
24.4b |
|
qGL2P-GW5+-qGS10B
|
5 |
5.84a |
2.86c |
1.96c |
2.05a |
23.4bc |
|
qGL2P-GW5+-qGS10P
|
14 |
5.61b |
2.75d |
1.87d |
2.04a |
20.1d |
Table 7Effects of qGL2, GW5, and qGS10 allele combinations on the phenotype of yield-related traits.
Table 7
|
Allele combinationz
|
n |
HDy
(DAS) |
CL
(cm) |
PL
(cm) |
PN |
NS |
TGW
(g) |
RRG
(%) |
BRR
(%) |
Yield
(g/plant) |
|
qGL2B-gw5--qGS10B
|
11 |
109abx
|
75bc |
20.3ab |
11.3a |
142ab |
24.0b |
72.4bc |
80.0bc |
28.4ab |
|
qGL2B-gw5--qGS10P
|
14 |
100bc |
72c |
19.6b |
11.2a |
118b |
22.4c |
86.3a |
80.6ab |
23.5cd |
|
qGL2B-GW5+-qGS10B
|
11 |
104abc |
77bc |
20.1ab |
11.1a |
150a |
22.2c |
73.7bc |
79.1bcd |
26.6bc |
|
qGL2B-GW5+-qGS10P
|
15 |
95c |
72c |
20.5ab |
11.2a |
119b |
20.3d |
72.9bc |
78.1d |
19.5d |
|
qGL2P-gw5--qGS10B
|
18 |
108ab |
81ab |
21.3a |
10.3a |
152a |
26.5a |
85.3a |
80.8ab |
28.5ab |
|
qGL2P-gw5--qGS10P
|
6 |
113a |
84a |
21.1a |
10.8a |
167a |
24.4b |
87.1a |
82.1a |
31.7a |
|
qGL2P-GW5+-qGS10B
|
5 |
100bc |
80ab |
21.3a |
10.9a |
159a |
23.4bc |
68.4c |
79.4bcd |
24.5bc |
|
qGL2P-GW5+-qGS10P
|
14 |
105ab |
78abc |
20.1ab |
10.5a |
158a |
20.1d |
80.5ab |
78.5cd |
23.4cd |
References
- 1. Ashfaq H, Rani R, Perveen N, Babar AD, Maqsood U, Asif M, Steele KA, Arif M. 2023. KASP mapping of QTLs for yield components using a RIL population in Basmati rice (Oryza sativa L.). Euphytica 219: 79
- 2. Calingacion M, Laborte A, Nelson A, Resurreccion A, Concepcion JC, Daygon VD, Mumm R, Reinke R, Dipti S, Bassinello PZ, Manful J, Sophany S, Lara KC, Bao J, Xie L, Loaiza K, EI A, Gayin J, Sharma N, Rajeswari S, Manonmani S, Rani NS, Kora S, Indrasari SD, Habibi F, Hosseini M, Tavasoli F, Suzuki K, Umemoto T, Boualaphanh C, Lee HH, Hung YP, Ramli A, Aung PP, Ahmad R, Wattoo JI, Bandonill EB, Romero M, Brites CM, Hafeel R, Lur HS, Cheaupun K, Jongdee S, Blanco P, Bryant R, Lang NT, Hall RD, Fitzgerald M. 2014. Diversity of global rice markets and the science required for consumer-targeted rice breeding. PloS One 9: e85106.
- 3. Chen E, Hou Q, Liu K, Gu Z, Dai B, Wang A, Feng Q, Zhao Y, Zhou C, Zhu J. 2023. Armadillo repeat only protein GS10 negatively regulates brassinosteroid signaling to control rice grain size. Plant Physio 192: 967-981.
- 4. Cheon KS, Jeong YM, Lee YY, Oh J, Kang DY, Oh HJ, Kim SL, Kim NH, Lee EY, Baek JH. 2019. Kompetitive allele-specific PCR marker development and quantitative trait locus mapping for bakanae disease resistance in Korean japonica rice varieties. Plant Breed Biotech 7: 208-219.
- 5. Cui S, Song P, Wang C, Chen S, Hao B, Xu Z, Cai L, Chen X, Zhu S, Gan X. 2024. The RNA binding protein EHD6 recruits the m6A reader YTH07 and sequesters OsCOL4 mRNA into phase-separated ribonucleoprotein condensates to promote rice flowering. Mol Plant. 17: 935-954.
- 6. Fan Y, Li Y. 2019. Molecular, cellular and Yin-Yang regulation of grain size and number in rice. Mol Breed 39: 163
- 7. Fujino K, Obara M, Ikegaya T. 2019. Establishment of adaptability to the northern-limit of rice production. Mol Genet Genomics 294: 729-737.
- 8. Garris A, Tai T, Coburn J, Kresovich S, McCouch S. 2005. Genetic structure and diversity in Oryza sativa L. Genetics 169: 1631-1638.
- 9. Glaszmann J. 1987. Isozymes and classification of Asian rice varieties. Theor Appl Genet 74: 21-30.
- 10. Juliano B. 1979. The chemical basis of rice grain quality. In: Proceedings of workshop on chemical aspects of rice grain quality. IRRI, Philippines. pp. 69-90.
- 11. Kim SR, Torollo G, Yoon MR, Kwak JE, Lee CK, Prahalada G, Choi IR, Yeo US, Jeong OY, Jena KK, Lee JS. 2018. Loss-of-function alleles of heading date 1 (Hd1) are associated with adaptation of temperate japonica rice plants to the tropical region. Front Plant Sci 9: 1827
- 12. Kim Y. 2010. Current status and future prospects of development of rice varieties. Food Preserv Proc Indus 9: 75-85.
- 13. Kovach MJ, Sweeney MT, McCouch SR. 2007. New insights into the history of rice domestication. Trends Genet 23: 578-587.
- 14. Kwon YH, Lee JY, Choi JS, Lee SM, Kim DJ, Kim PJ, Chae HG, Cha JK, Park HJ, Kang JW, Kim TH, Chae HG, Kabange NR, Oh KW, Kim PJ, Kwak YS, Lee JH, Ryu CM. 2023. Loss-of-function gs3 allele decreases methane emission and increases grain yield in rice. Nat Clim Change 13: 1329-1333.
- 15. Lee CM, Lee KM, Baek MK, Kim WJ, Suh JP, Jeong OY, Cho YC, Park HS, Kim SM. 2020. Characterization of traits related to grain shape in Korean rice varieties. Korean J Crop Sci 65: 199-213.
- 16. Lee CW, Cheon KS, Shin YJ, Oh HJ, Jeong YM, Jang H, Park YC, Kim KY, Cho HC, Won YJ, Baek JH, Cha YS, Kim SL, Kim KH, Ji HS. 2022. Development and application of a target capture sequencing SNP-genotyping platform in rice. Genes 13: 794
- 17. Lee SY, Jeung JU, Mo YJ. 2024. Allelic combinations of Hd1, Hd16, and Ghd7 exhibit pleiotropic effects on agronomic traits in rice. G3: Genes, Genomes, Genetics 14: jkad300.
- 18. Liu J, Chen J, Zheng X, Wu F, Lin Q, Heng Y, Tian P, Cheng Z, Yu X, Zhou K. 2017. GW5 acts in the brassinosteroid signalling pathway to regulate grain width and weight in rice. Nat Plant 3: 1-7.
- 19. Lu H, Redus MA, Coburn JR, Rutger JN, McCouch SR, Tai TH. 2005. Population structure and breeding patterns of 145 US rice cultivars based on SSR marker analysis. Crop Sci 45: 66-76.
- 20. Lu Y, Chuan M, Wang H, Chen R, Tao T, Zhou Y, Xu Y, Li P, Yao Y, Xu C. 2022. Genetic and molecular factors in determining grain number per panicle of rice. Front Plant Sci 13: 964246.
- 21. Lyu J, Wang D, Duan P, Liu Y, Huang K, Zeng D, Zhang L, Dong G, Li Y, Xu R, Zhang B, Huang X, Li N, Wang Y, Qian Q, Li Y. 2020. Control of grain size and weight by the GSK2-LARGE1/OML4 pathway in rice. Plant Cell 32: 1905-1918.
- 22. Meng L, Li H, Zhang L, Wang J. 2015. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. T Crop J 3: 269-283.
- 23. Mo YJ, Lee CM, Park HM, Ha SK, Kim MJ, Kwak JE, Lee HS, Lee JH, Jeung JU. 2021. Hd1 allele types and their associations with major agronomic traits in Korean rice cultivars. Plants 10: 2408
- 24. Nam JK, Park HS, Baek MK, Cho YC, Kim WJ, Kim JJ, Kim B-K, Kim KY, Shin WC, Ko JC, Lee GM, Park SG, Lee CM, Kim CS, Suh JP, Lee JH. 2019. Bacterial blight-resistant medium maturing rice cultivar 'Haepum' with high grain quality. Korean J Breed Sci 51: 222-233.
- 25. Park HS, Baek MG, Nam JK, Shin WC, Lee GM, Park SG, Lee CM, Kim CS, Cho YC. 2018. Development and characterization of japonica rice line with long and spindle- shaped grain. Korean J Breed Sci 50: 116-130.
- 26. Park HS, Baek MK, Lee CM, Seo JH, Park SH, Kang KM, Park JR, Jin MA, Jeong OY, Jeung JU, Kim KY. 2023. High grain quality mid-late maturing rice cultivar 'Namchan' with high yield performance under low-nitrogen fertilizer conditions. Korean J Breed Sci 55: 389-399.
- 27. Park HS, Baek MK, Nam JK, Shin WC, Jeong JM, Lee GM, Park SG, Kim CS, Cho YC, Kim BK. 2017. Development and characterization of breeding materials with diverse grain size and shape in japonica. rice Korean J Breed Sci 49: 369-389.
- 28. Park HS, Ha KY, Kim KY, Kim WJ, Nam JK, Baek MK, Kim JJ, Jeong JM, Cho YC, Lee JH, Kim BK, Ahn SN. 2015. Development of high-yielding rice lines and analysis of panicle and yield-related traits using doubled haploid lines derived from the cross between Deuraechan and Boramchan, high-yielding japonica rice cultivars in Korea. Korean J Breed Sci 47: 384-402.
- 29. Park HS, Seo JH, Lee CM, Park SH, Lee KM, Park JR, Jeong OY. 2024a. Quantitative trait loci analysis of quality-related traits using recombinant inbred lines derived from a cross between 'Chamdongjin'and 'Younghojinmi'. Korean J Breed Sci 56: 395-415.
- 30. Park HS, Seo JH, Park SH, Park JR, Lee CM, Jin MA, Jeong OY. 2024b. QTL analysis for yield-related traits using the recombinant inbred lines derived from a cross between 'Chamdongjin' and 'Younghojinmi'. Korean J Breed Sci 56: 31-51.
- 31. Park HS, Suh JP, Baek MK, Lee CM, Kim WJ, Lee GM, Kim SM, Kim CS, Cho YC. 2020. Characterization of yield and panicle-related traits of early maturing rice varieties by cultivation times in the Honam plain area of Korea. Korean J Breed Sci 52: 115-130.
- 32. Park JR, Lee CM, Baek MK, An JH, Seo JH, Hong HC, Jeong OY, Park HS. 2022. Characterization of grain-related traits and pasting and texture properties of United State rice varieties in Korea. Korean J Breed Sci 54: 81-97.
- 33. Park JR, Seo JH, Park SH, Jin MA, Jeong OY, Park HS. 2023. Identification of potential QTLs related to grain size in rice. Plants 12: 1766
- 34. Sasaki A, Swapan D, Ishiyama K, Saito T. 2002. A mutant gibberellin-synthesis gene in rice. Nature 416: 701-702.
- 35. Seo JH, Reflinur , Kim SH, Woo MO, Koh HJ. 2014. Identification of quantitative trait loci for agronomic traits in two rice populations derived from a cross with a wide compatability line. Plant Breed Biotech 2: 231-246.
- 36. Sha H, Liu H, Zhao G, Han Z, Chang H, Wang J, Zheng H, Zhang J, Yu Y, Liu Y, Zou D, Nie S, Fang J. 2022. Elite sd1 alleles in japonica rice and their breeding applications in northeast China. Crop J 10: 224-233.
- 37. Singh VJ, Bhowmick PK, Vinod KK, Krishnan SG, Nandakumar S, Kumar A, Kumar M, Shekhawat S, Dixit BK, Malik A. 2022. Population structure of a worldwide collection of tropical japonica rice indicates limited geographic differentiation and shows promising genetic variability associated with new plant type. Genes 13: 484
- 38. Sun K, Zong W, Xiao D, Wu Z, Guo X, Li F, Song Y, Li S, Wei G, Hao Y. 2023. Effects of the core heading date genes Hd1, Ghd7, DTH8, and PRR37 on yield-related traits in rice. Theor Appl Genet 136: 227
- 39. Takahashi Y, Shomura A, Sasaki T, Yano M. 2001. Hd6, a rice quantitative trait locus involved in photoperiod sensitivity, encodes the α subunit of protein kinase CK2. Proc Natl Acad Sci USA 98: 7922-7927.
- 40. Vaughan DA, Lu BR, Tomooka N. 2008. The evolving story of rice evolution. Plant Sci 174: 394-408.
- 41. Wang M, Huang L, Kou Y, Li D, Hu W, Fan D, Cheng S, Yang Y, Zhang Z. 2023. Differentiation of morphological traits and genome-wide expression patterns between rice subspecies indica and japonica. Genes 14: 1971
- 42. Wang W, Mauleon R, Hu Z, Chebotarov D, Tai S, Wu Z, Li M, Zheng T, Fuentes RR, Zhang F. 2018. Genomic variation in 3,010 diverse accessions of Asian cultivated rice. Nature 557: 43-49.
- 43. Yano M, Katayose Y, Ashikari M, Yamanouchi U, Monna L, Fuse T, Baba T, Yamamoto K, Umehara Y, Nagamura Y. 2000. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. The Plant Cell 12: 2473-2483.
- 44. Zhan P, Wei X, Xiao Z, Wang X, Ma S, Lin S, Li F, Bu S, Liu Z, Zhu H. 2021. GW10, a member of P450 subfamily regulates grain size and grain number in rice. Theor Appl Genet 134: 3941-3950.
- 45. Zhang ZH, Wang K, Guo L, Zhu YJ, Fan YY, Cheng SH, Zhuang JY. 2012. Pleiotropism of the photoperiod-insensitive allele of Hd1 on heading date, plant height and yield traits in rice. PloS One 7: e52538.