적요
Wheat is a fundamental staple crop worldwide, contributing significantly to global food security due to its versatility and nutritional value. However, gluten proteins, including gliadins and glutenins, have been implicated in various health problems, such as celiac disease, non-celiac gluten sensitivity, and wheat allergies. These disorders affect a wide variety of people globally, creating demand for wheat varieties that balance high-end-use quality with reduced immunogenic potential. This review examines the molecular and genetic mechanisms that regulate gluten protein synthesis, highlighting recent advances in genomic and mutagenic approaches aimed at modifying gluten proteins to enhance the health and quality traits of wheat. Technologies such as RNAi and CRISPR/Cas9 offer promising avenues for reducing wheat immunogenicity without compromising its functional properties in food production. This study also examines the challenges and prospects of utilizing these genetic tools to develop wheat varieties that achieve the dual objectives of enhanced health outcomes and high product quality.
서언
밀은 벼, 옥수수와 더불어 전 세계적으로 널리 재배되고 소비되는 주요 식량작물로, 전 세계 인구가 소비하는 칼로리와 단백질의 약 20%를 담당하고 있고, 주요 에너지원이자 필수 미량원소 및 섬유질 공급원으로서 다양한 활용성을 가진 중요한 주식으로 자리 잡고 있다(
Gupta et al. 2008,
Shewry & Hey 2015). 전 세계 밀 생산량은 1969년 3억톤에서 2023년에는 7억 9,828만 톤으로 증가하였으며, 그 수요가 증가함에 따라 밀 생산은 꾸준히 증가해왔다(
FAOSTAT 2022,
KREI 2024,
Wieser et al. 2022). 향후 인구 증가에 따른 식량 수요가 2050년까지 35~56% 증가할 것으로 예상되며, 밀은 고단백 식품원으로 영양 수요를 충족하는 데 중요한 역할을 하므로 식량안보의 초석으로 계속 남을 것으로 예상된다(
van Dijk et al. 2021). 또한 밀은 우리의 식탁에서 하루도 빠질 수 없는 가장 다양하게 활용되고 있는 우리나라 제2의 주곡 작물이다. 국내 밀 수요량은 220여만 톤으로 국내 밀 자급률은 약 1%에 불과해 대부분 수입에 의존하는 실정이다. 이러한 상황은 식량안보 측면에서 자급률 증대, 고부가가치 품종 개발 등 특단의 대책이 필요한 상황이다.
주요 밀 종은 육배체인
Triticum aestivum (AABBDD, 2n=6x=42)으로 전 세계적으로 재배되며 흔히 “보통계밀” 또는 “빵밀”로 알려져 있다(
Shewry 2009). 전 세계 밀 생산에는 지중해 유역의 건조한 환경과 이와 유사한 기후에 적합한 사배체 종인
T. turgidum subsp.
durum (AABB, 2n=4x=28)도 포함된다. 주로 파스타 생산에 사용되는 이 종은 일반적으로 “듀럼밀” 또는 “파스타밀”이라 불린다. 반면 종종 문화적 선호도 또는 건강식품에 대한 수요로 인해 제한된 양만 재배되는 밀 종도 있다. 여기에는 이배체 종인 Einkorn (
T. monococcum subsp.
monococcum, AA, 2n=2x=14), 사배체 종인 엠머밀(
T. turgidum subsp.
dicoccum, AABB, 2n=28), 그리고 육배체 재배 밀인 스펠트밀(
T. aestivum subsp.
spelta, AABBDD, 2n=6x=42)이 포함된다.
글루텐은 밀에 존재하는 단백질 복합체로 밀가루 특유의 쫄깃하고 찰진 식감을 만들어주고 빵을 부풀게 하는 성분으로서 반죽에 독특한 점탄성(viscoelasticity) 특성을 부여하여 빵에서 국수에 이르기까지 다양한 식품의 생산을 용이하게 한다(
Khan 2019). 밀 글루텐 단백질은 글루테닌과 글리아딘으로 구성되어 있으며, 밀가루에 물을 첨가하여 반죽하면 글루텐 복합체(gluten complex)가 형성된다(
Delcour et al. 2012,
Singh & MacRitchie 2001). 글루테닌과 글리아딘의 유전자형(allelic composition)이 가공적성(빵, 면, 과자 등)에 많은 영향을 미치며, 용도별 최고 품질의 밀 육성에 중요한 요인이 되고 있다(
Filip et al. 2023).
전 세계적으로 많은 사람들이 밀 관련 질환(wheat-related disorders)으로 고통받고 있다. 대표적인 밀 관련 질환으로 밀 알레르기(wheat allergy), 셀리악병(celiac disease), 비셀리악 글루텐 민감증(non-celiac gluten sensitivity, NCGS)이 있으며, 이들 질환의 주요 항원 단백질이 결손된 밀을 개발하는 연구가 전 세계적으로 수행되고 있다(
Gil-Humanes et al. 2010,
Lindfors et al. 2019). 셀리악병은 글루텐 펩타이드에 의해 유도되는 자가면역 질환으로 전 세계 인구의 약 1%에 영향을 미치고 있다(
Marchioni Beery & Birk 2015,
Taraghikhah et al. 2020). 또 다른 글루텐 관련 질환인 비셀리악 글루텐 민감증은 성인 여성에게 더 흔하며 일반 인구의 약 0.6~13%에 달한다. 이러한 질환의 병리학적 원인은 아직 잘 이해되지 않고 있으며, 현재로서는 글루텐-프리 식단을 엄격히 유지하는 것이 글루텐 관련 질환에 대한 유일한 치료법이다. 또한 밀-의존성 운동-유발성 아나필락시스(wheat-dependent exercise-induced anaphylaxis, WDEIA)를 포함한 밀 알레르기도 많은 사람에게 추가적인 밀의 식이 제한을 가하고 있다(
Sharma et al. 2020). 이러한 질환의 주요 메커니즘은 특정 글루텐 펩타이드에 대한 신체의 면역 반응과 관련이 있다. 이 외에도 포진성 피부염과 글루텐 운동실조증도 중요한 글루텐 관련 질환으로 알려져 있는데, 포진성 피부염은 심한 가려움증을 특징으로 하는 만성 수포성 피부 질환이며 글루텐 민감증과 직접적인 관련이 있고, 글루텐 운동실조증은 자가면역 질환으로서 소뇌의 뉴런을 표적으로 삼아 균형과 조정 능력을 저하시킨다(
Sapone et al. 2012). 이러한 글루텐 관련 질환 문제가 증가함에 따라 글루텐-프리 식단은 글루텐 관련 질환에 대한 필수적인 치료법으로 대두되고 있다.
한국, 일본, 중국 등의 동아시아에서는 밀 관련 질환에 대한 보고가 드물지만, 이는 유전적 감수성 차이도 있지만 환경적 요인에서 기인한 것으로 추정된다(
Cummins & Roberts-Thomson 2009,
Ganbaatar et al. 2025). 인도네시아, 베트남, 필리핀과 같은 동남아시아에서는 셀리악병이 인구의 5% 이상의 유병률을 나타낸다고 보고되기도 하지만, 주식이 쌀이기 때문에 밀 소비량이 낮아 유병률이 드문 것으로 예측된다(
Yachha 2006). 즉 아직은 아시아 국가들이 밀 관련 질환과 연관성이 낮아 보일 수 있지만, 식습관이나 환경조건에 따라 밀 관련 질환의 위험성이 증가할 가능성이 잠재되어 있다. 또한 아시아에서는 밀 관련 질환에 대한 선별검사가 활발히 이루어지지 않아 실제 유병률이 과소평가 되었을 가능성도 있다(
Makharia 2015). 따라서 서구권 못지않게 한국을 포함한 아시아에서도 밀 관련 질환에 대해 이해하고 관련 건강 문제에 대비하기 위한 연구가 필요하다. 비셀리악 글루텐 민감증과 밀 알레르기에 대한 연구는 서구권을 중심으로 진행되어 왔으나, 최근 아시아에서도 관심이 증가하고 있다(
Gil-Humanes et al. 2010,
Lindfors et al. 2019). 한국에서 수행된 한 연구에서는 글루텐 섭취 후 소화기 증상과 신경학적 증상을 호소하는 환자들이 있었으나 셀리악병 진단 기준을 충족하지 않는 경우가 다수 보고되었다(
Cha & Kim 2020,
Lee & Lee 2019). 이는 비셀리악 글루텐 민감증의 존재 가능성을 시사하며, 이와 관련하여 보다 정밀한 연구가 필요한 실정이다. 한국과 일본에서는 밀 알레르기 관련 연구가 활발히 이루어지고 있으며, 특히 소아의 면역 반응과 아나필락시스 발생률에 대한 연구가 진행되었다(
Jeong et al. 2017,
Koike et al. 2018,
Sato et al. 2015). 또한 밀가루를 포함한 가공식품 섭취와 관련된 알레르기 반응이 점점 증가하고 있다고 보고되었다.
글루텐 관련 건강 문제를 해결하기 위한 밀 육종에 있어서, 질병에 대한 감수성을 줄이는 동시에 밀 품질을 개선하는 목표가 종종 충돌하면서 밀 육종의 어려움이 드러나고 있다. 글루텐이 관련 질환의 면역원성(immunogenic potential)에 연관됨과 동시에 제빵 품질 유지 기능을 가질 뿐 아니라, 밀과 글루텐 단백질의 복잡한 유전적 구조로 인해 전통적인 육종은 더욱 어려움을 겪고 있다(
Khan 2019). 그러나 최근 분자 육종 기술의 발전으로 이러한 복잡성을 해결할 가능성이 제시되고 있고, 글루텐 구성요소를 보다 정밀하게 수정하여 건강 문제와 가공 특성을 동시에 개선할 수 있는 기회가 제공되었다(
Wang et al. 2020). 또한 글루텐 단백질의 조절에 관한 최근 연구는 밀 품질과 다양한 가공 적합성에 대한 이해를 크게 향상시켰고, 유전체학 및 분자생물학의 발전으로 글루텐 단백질 구성의 정밀한 조작이 가능해졌으며, 이를 통해 특정 품질 특성을 갖춘 밀 품종을 개발할 수 있게 되었다(
Walkowiak et al. 2020). 본 총설에서는 글루텐 단백질에 대한 일반적인 소개와 전통 육종 및 생명공학 기법을 이용하여 밀 관련 질환의 면역원성을 줄이기 위한 최신 연구 동향에 대해 소개할 것이다.
글루텐 단백질의 구조와 기능
글루텐 단백질의 분류, 구조, 기능 및 유전
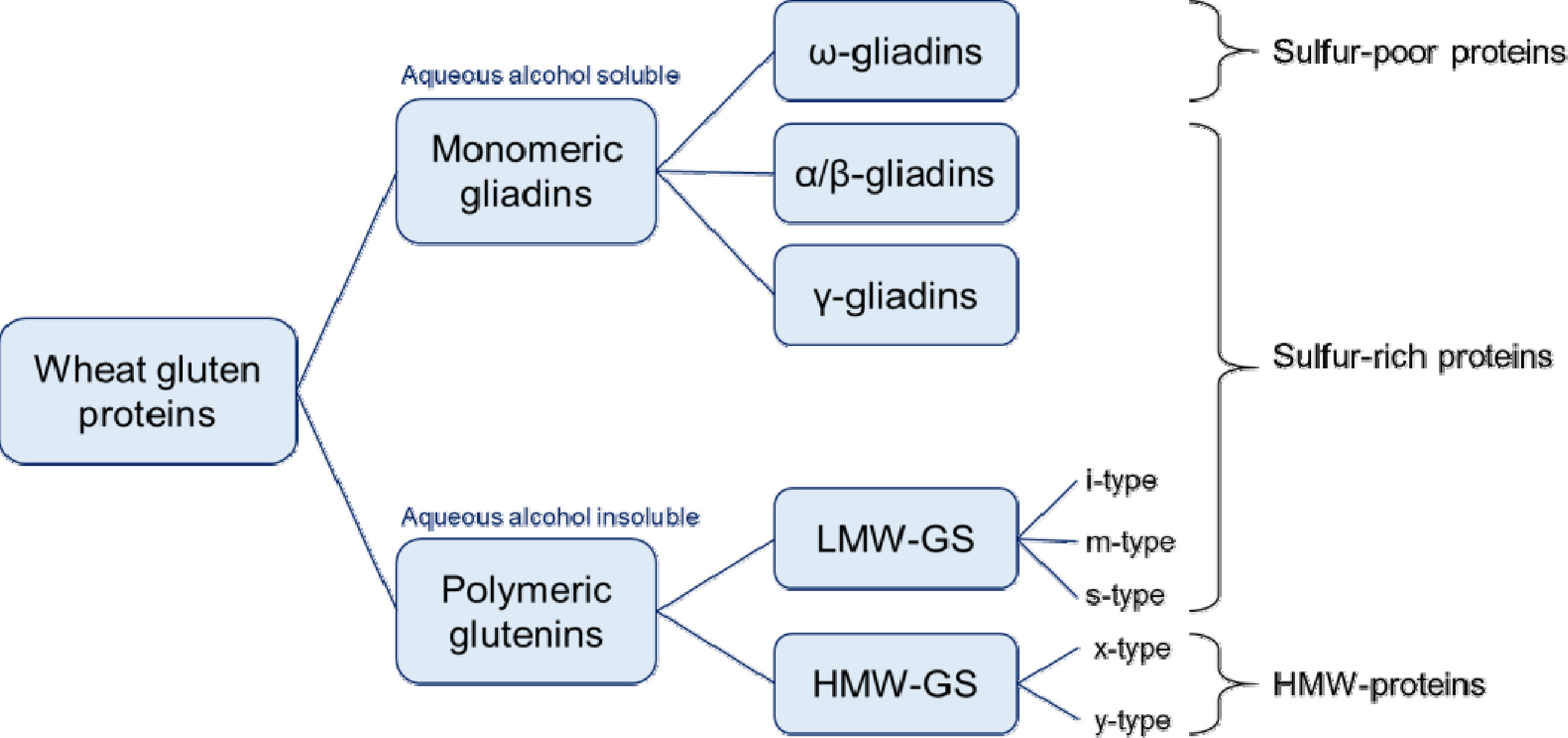
밀의 상업적 가치는 다른 곡물들과 달리 글루텐 단백질에 의해 기인하며, 밀 반죽에 점탄성(viscoelasticity)을 부여하여 빵, 면, 쿠키, 케이크 등과 같은 다양한 음식을 인류에게 제공해 왔다. 글루텐 단백질은 알코올(60-80% aqueous ethanol)에 녹는 단량체의 글리아딘(gliadin)과 녹지 않는 다량체의 글루테닌(glutenin)으로 분류된다(
Fig. 1) (
Shewry & Halford 2002,
Shewry & Tatham 1997). 일반적으로 글루테닌은 밀 반죽에 탄성(elasticity)과 힘(strength)을 부여하고 글리아딘은 점성(viscosity)과 신장성(extensibility)을 부여한다(
Wieser 2007). 글루테닌은 분자량에 따라 고분자 글루테닌 서브유닛(high-molecular-weight glutenin subunit, HMW-GS; MW 67-88 kDa)과 저분자 글루테닌 서브유닛(low-molecular- weight glutenin subunit, LMW-GS; MW 32-35 kDa)으로 구성되며, 이들은 분자간 및 분자내 이황화 결합(inter- and intra-chain disulfide bond)에 의해 밀 반죽에서 거대고분자 복합체(macropolymer complex)를 형성한다(
Wieser et al. 2023). 이들 글루테닌은 시스테인 잔기에 존재하는 황이 풍부하여 S-rich prolamin이라고 불린다(
Shewry 2019).
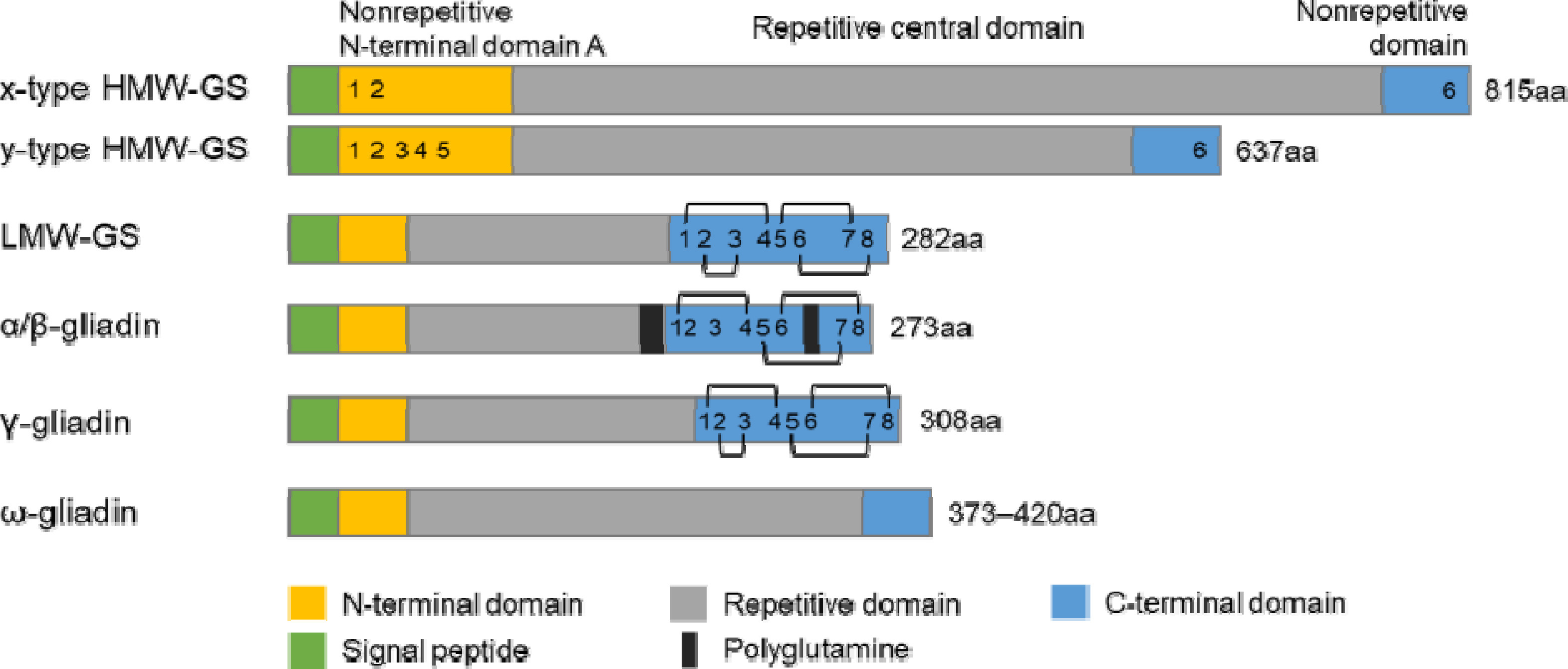
HMW-GS는 총 글루텐 함량에 약 5-10% 정도 차지하지만, 글루테닌 중합체의 형성을 촉진하기 때문에 밀 반죽의 탄성, 힘, 제빵 가공성에 중요한 역할을 한다(
Don et al. 2006,
Köhler et al. 1991, 1993,
Lee et al. 2011,
Payne et al. 1987,
Shewry et al. 1992,
Shewry & Lafiandra 2022,
Tao et al. 1992 ). 이들의 구조는 시그널 펩타이드, N-말단, 반복 도메인, C-말단으로 이루어지며, 분자간 및 분자내 이황화 결합을 형성하는 시스테인 잔기의 위치, SDS-PAGE 이동성(mobility)과 구성 아미노산 서열에 따라 x-type HMW-GS (트리펩타이드 모티프, GQQ; 헥사펩타이드 모티프, PGQGQQ; 노나펩타이드 모티프, GYYPTSPQQ)와 y-type HMW-GS (헥사펩타이드 모티프, PGQGQQ; 노나펩타이드 모티프, GYYPTSLQQ)로 나뉜다(
Fig. 2) (
Anderson & Greene 1989,
Anjum et al. 2007,
Li et al. 2021,
Shewry et al. 2003,
Tatham et al. 1990). HMW-GS는 밀 염색체 1A, 1B, 1D의 장완(long arm)에 존재하는
Glu-A1,
Glu-B1,
Glu-D1 유전자좌에 의해 암호화되며(
Table 1), 각각의 유전자좌는 x-type HMW-GS와 y-type HMW-GS를 암호화하여 총 6개의 HMW-GS 유전자(allele)를 가지게 된다(
Gu et al. 2006,
Li et al. 2021,
Payne et al. 1980,
Shewry & Tatham 1997). 일반적으로 gene silencing으로 밀 한 품종에 3-5개의 HMW-GS가 발현되며, 이들의 구성 성분과 함량은 밀 반죽 특성 및 제빵 가공성에 많은 영향을 미친다고 알려져 있다(
Ng & Bushuk 1988,
Yahata et al. 2006). 특히
Ng & Bushuk (1988)은 HMW-GS의 특정 구성성분이 제빵 적성에 얼마나 관여하는지를 수식화하였다.
LMW-GS는 총 글루텐 함량에서 약 40%로 구성되며 분자량은 30-50 kDa 정도이고, 밀가루 반죽의 탄성과 신장성에 관여한다고 알려져 있다(
Wrigley 1996). 보통 LMW-GS는 시그널 펩타이드, N-말단, 반복 도메인, C-말단으로 구성되며, 시그널 펩타이드를 제거한 첫 아미노산 잔기에 따라 i-type (isoleucine, 이소루신), m-type (methionine, 메티오닌), s-type (serine, 세린) LMW-GS로 분류된다(
Fig. 1) (
D′Ovidio & Masci 2004). LMW-GS는 밀 염색체 1A, 1B, 1D의 단완(short arm)에 존재하는
Glu-A3,
Glu-B3,
Glu-D3 유전자좌에 의해 암호화되며(
Table 1), 이들의 함량 및 유전적 변이(allelic variation)가 밀 반죽 물성 및 가공성에 영향을 미친다고 알려져 있다(
Dong et al. 2010,
Singh & Shepherd 1988).
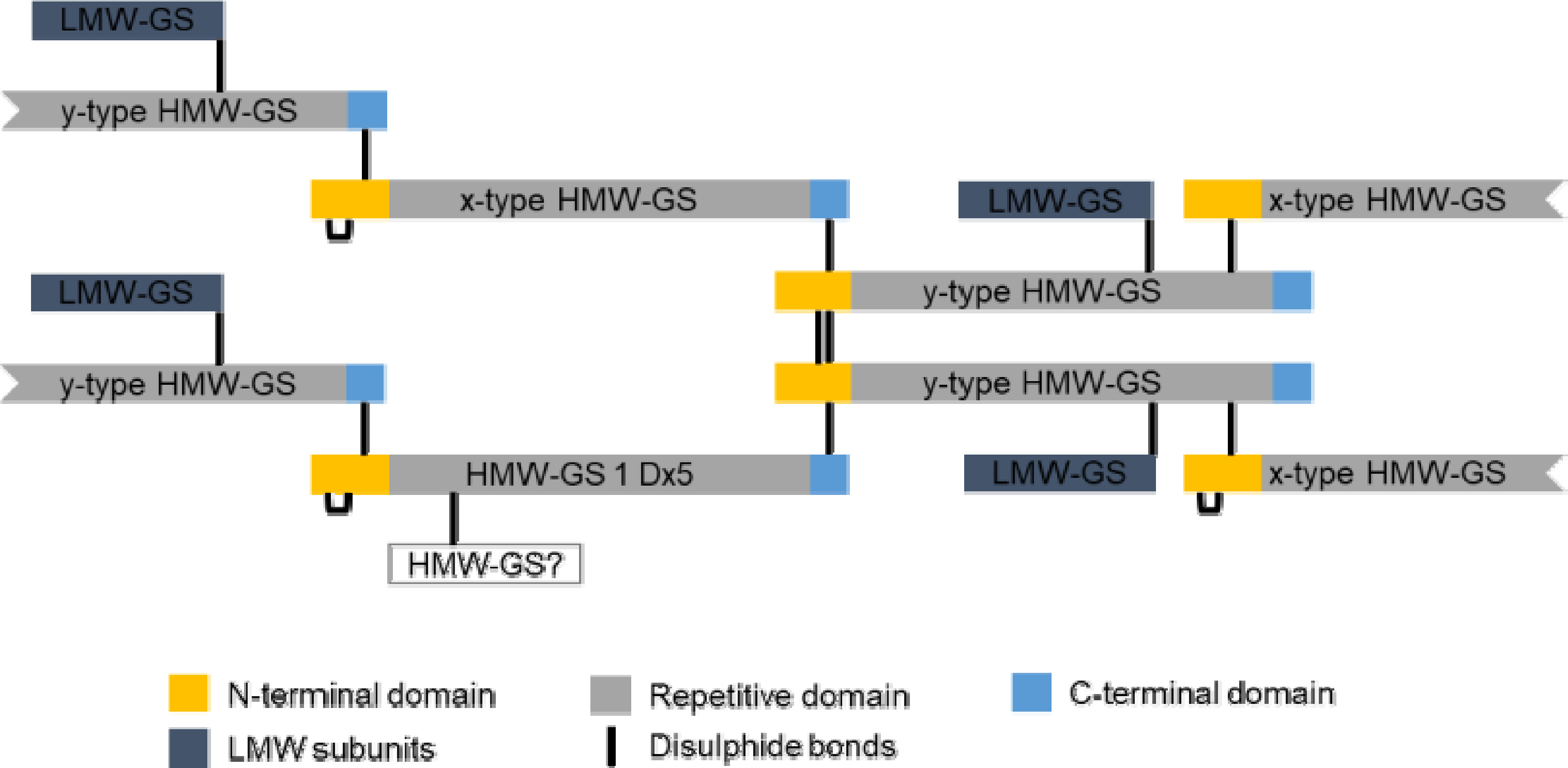
밀 반죽에서 글루텐은 거대고분자 복합체로 존재한다.
Fig. 3에서처럼 HMW-GS와 LMW-GS는 이황화 결합을 통해 거대고분자 복합체의 골격(backbone)을 형성하고 단량체 글리아딘 단백질들은 골격 중간에서 윤활유와 같이 점성을 부여하는 역할을 한다(
Anjum et al. 2007,
Belton 1999,
Shewry et al. 2003,
Wrigley 1996). 밀 반죽을 효모와 함께 발효시키면 발효 과정 중에 발생하는 이산화탄소를 거대고분자 복합체가 포획하여 빵이 부풀게 된다(
Cornish et al. 2006,
Shewry et al. 1995). 여러 연구에 따르면 HMW-GS와 LMW-GS 특정 유전자좌의 조성 조합(allelic combination)이 밀가루의 제빵 품질에 중요한 요소로서 강력분(빵), 중력분(면), 박력분(쿠키, 케이크)과 상관관계가 있다고 밝혀졌다(
Altpeter et al. 1996,
Ikeda et al. 2014). 특히 HMW-GS의
Glu-D1d (1Dx5+1Dy10) 또는
Glu-B1a (1Bx7
OE+1By8*), LMW-GS의
Glu-A3g,
Glu-B3b,
Glu-D3b의 유전자좌 조성 조합이 빵 가공성의 강력분 또는 초강력분과 관련된다고 알려져 있다(
Butow et al. 2003,
Farahani et al. 2023,
Gao et al. 2018,
Karaduman et al. 2022,
Radovanovic et al. 2002,
Sissons et al. 2005). 이러한 특성을 기반으로 ‘Glu1-score’라는 평가 시스템이 개발되었으며, 이 시스템은 품질 평가에 따라 개별 소단위체에 숫자 등급을 할당하여 밀의 제빵 품질을 평가하는 데 도움이 된다(
Payne et al. 1987). Glu1-score가 높을수록 글루텐이 강해져 반죽의 탄력성과 제빵 품질이 더 좋아지는 것을 의미한다.
글리아딘 단백질은 전체 글루텐 함량 중 약 40%를 차지하며, 밀 반죽에 점성과 신장성을 부여한다. 이들의 분자량은 28-75 kDa 범위이고, A-PAGE (acid-polyacrylamide gel electrophoresis)의 이동성과 이들의 일차구조에 따라 ω-글리아딘(ω5, ω1,2-글리아딘 포함), α/β-글리아딘, γ-글리아딘으로 분류된다(
Figs. 1,
2) (
Shewry 2019,
Wieser 2007). α/β- 및 γ-글리아딘의 단백질 구조는 시그널 펩타이드, 짧은 N-말단, 다양한 크기의 반복 도메인, 긴 C-말단으로 구성되며, C-말단에 존재하는 시스테인 잔기들이 분자내 이황화 결합(intra-chain disulfide bond)을 형성한다(
Urade et al. 2018,
Wieser et al. 2020,
Zhang et al. 2022). ω-글리아딘의 구조는 시그널 펩타이드, 짧은 N-말단, 긴 반복 도메인, 짧은 C-말단으로 구성되며, 시스테인 잔기가 없어 이황화 결합을 형성하지 않는다(
Barak et al. 2015). 글리아딘은 밀 염색체 1번과 6번의 단완에 위치한
Gli-1과
Gli-2 유전자좌에 의해 암호화된다. ω1,2-글리아딘은 1번 염색체의 단완에 위치한
Gli-A1,
Gli-D1 유전자좌에 의해 암호화되고, ω5-글리아딘은
Gli-B1 유전자좌에 의해 암호화된다. γ-글리아딘은
Gli-A1,
Gli-B1,
Gli-D1 유전자좌에 의해 암호화되고, α/β-글리아딘은 6번 염색체의 단완에 위치한
Gli-A2,
Gli-B2,
Gli-D2 유전자좌에 의해 암호화된다(
Ferranti et al. 2007).
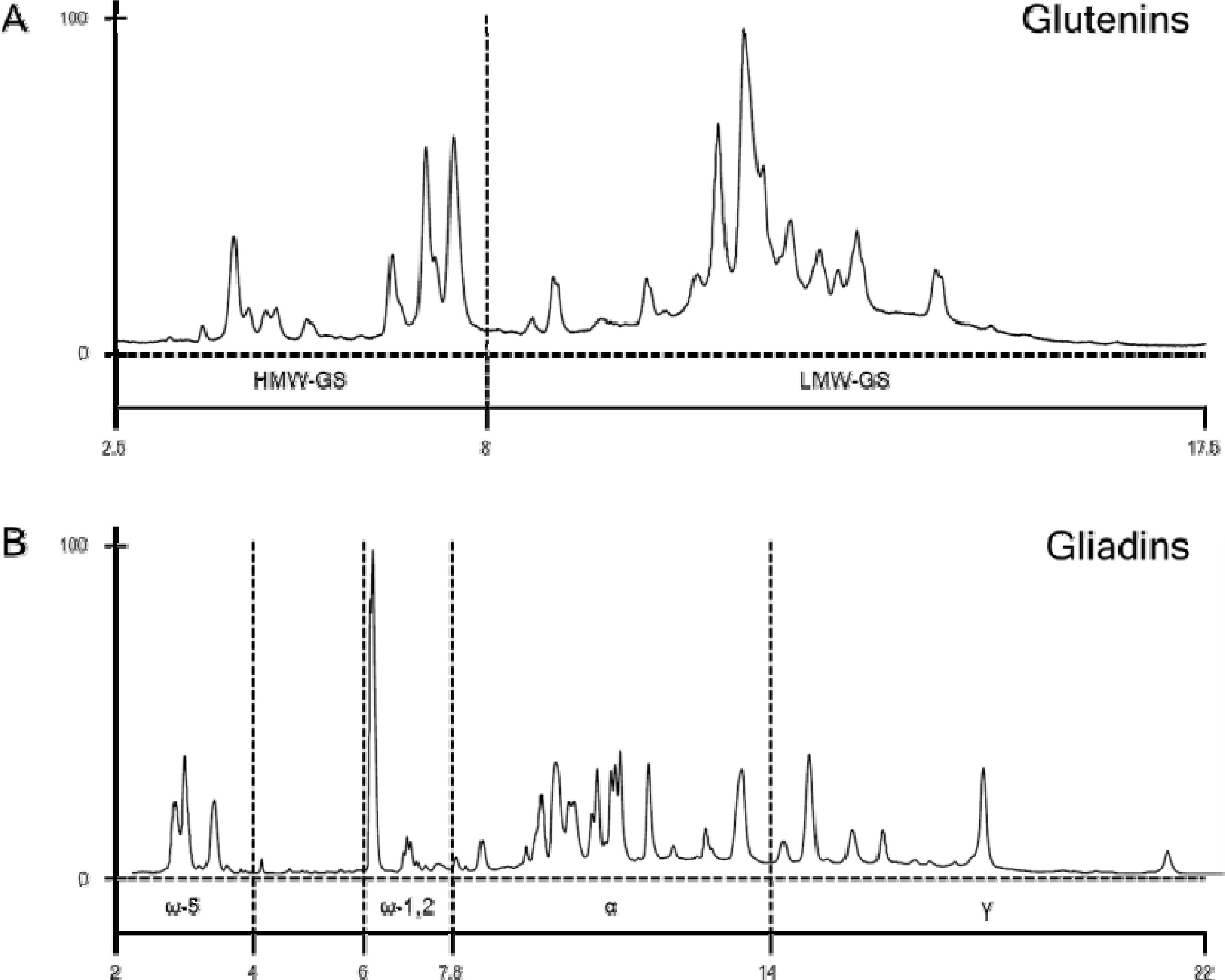
Fig. 4에 밀의 유전 육종 연구에 사용되는 레퍼런스 품종 Chinese Spring의 글루테닌과 글리아딘 분획의 프로파일을 나타내었다. 글루테닌 분획에는 HMW-GS 및 LMW-GS 부분, 글리아딘 분획에는 ω5-, ω1,2-, α-, γ-글리아딘 영역을 각각 표시하였다.
밀 글루텐은 셀리악병이나 글루텐 민감증이 있는 사람에게는 심각한 건강 문제를 야기할 수 있다. 다시 말해 글리아딘 및 특정 글루테닌 펩타이드에 의해 유발되는 면역반응은 심각한 위장 증상과 장기적인 건강 문제로 이어질 수 있으며(
Biesiekierski et al. 2013), 따라서 밀 글루텐의 세부적인 특성을 이해하는 것은 밀 글루텐 섭취와 관련된 건강 문제를 해결하는 데 필수적이다.
최근 연구에 따르면 글루텐의 반복적인 서열이 셀리악병을 유발하는 주요 원인으로 여겨지고 있다(
Shewry 2019). 셀리악병은 자가면역 유사 질환으로, 탈아미노 된 글루텐 펩타이드에 대한 CD4
+ T세포의 반응으로 인해 장 점막이 손상되는 질환이며, 해당 펩타이드는 T세포의 표면에 존재하는 일부 조직적합성항원(human leukocyte antigen, HLA) 분자에 의해 면역반응을 유발한다(
Chlubnová et al. 2023). 셀리악병과 연관된 항원결정기(epitope)는 밀 글루텐의 반복 도메인에 존재하며, 글리아딘과 글루테닌은 항원결정기를 포함하고 있다(
Sapone et al. 2012). 셀리악병 발병 기전에 관여하는 가장 주목할 만한 글루텐 항원결정기는 HLA-DQ2.5 및 HLA-DQ8와 관련이 있으며, HLA-DQ2.2는 소수의 경우에서 관련이 있는 것으로 나타났다(
Sollid et al. 2012).
WDEIA를 포함한 밀 알레르기는 면역글로불린 E (immunoglobulin E, IgE)-매개 과민반응으로 면역학적 기전을 수반하고, 두드러기에서 혈관부종, 호흡곤란, 저혈압, 쇼크에 이르기까지 다양한 아나필락시스(anaphylaxis) 반응이 특징이다(
Battais et al. 2005,
Scherf et al. 2016). WDEIA는 일반적으로 밀 기반 식품을 섭취한 후 신체 운동을 한 후에 발생하며, ω5-글리아딘이 주요 알레르겐(allergen)으로 보고되었다(
Altenbach et al. 2018,
Faihs et al. 2023). ω5-글리아딘의 항원결정기(IgE 결합 부위)는 QQX
1PX
2QQ (X
1은 L, F, S, I 중 하나이고 X
2는 Q, E, G 중 하나임)이다(
Battais et al. 2005,
Matsuo et al. 2004, 2005,
Morita et al. 2023,
Palosuo et al. 1999, 2001). 지금까지 알려진 WDEIA의 예방법은 글루텐이 없는 식단이며 특히 운동 전 밀 섭취를 엄격히 제한한다.
최근 Chinese Spring 품종에서 셀리악병과 WDEIA 관련 항원결정기를 포함하는 글리아딘과 글루테닌 단백질의 특성이 분석되었으며, 가장 면역원성과 독성이 높은 유형은 α-와 ω-글리아딘에 위치한 것으로 밝혀졌다(
Tables 2,
3) (
Altenbach et al. 2020a,
Huo et al. 2018). 대부분의 α-글리아딘 유전자는 셀리악병을 유발할 가능성이 있는 단백질을 암호화하며, 이들 중 일부는 3개의 항원결정기가 중첩된 사본 6개로 구성된 33-mer 펩타이드를 포함하고 있고, 이는 특히 강력한 T세포 자극제로 간주된다(
Schaart et al. 2020). Chinese Spring의 단백질체에 관한 최근 연구는 글루텐 단백질 유전자 중 총 42가지가 셀리악병 또는 WDEIA와 관련이 있다고 보고하였으며, 이 중에는 16개의 α-글리아딘, 11개의 γ-글리아딘, 7개의 ω-글리아딘, 8개의 LMW-GS가 포함된다(
Huo et al. 2018). 또한, 잘린 형태의 단백질을 암호화하는 두 ω-글리아딘 유전자(ω-A4, ω-D4)도 확인되었다. 이러한 연구 결과는 밀의 면역원성을 줄이기 위한 생명공학적 방법을 고안하는 데 매우 중요한 정보를 제공할 것이다. 특히 글루텐 단백질 유전자 발현 조절을 통해 셀리악병 및 WDEIA를 유발하는 항원결정기를 제거하거나 감소시켜 밀 기반 제품의 건강성을 향상시키는 데 기여할 수 있다.
밀의 가공적성 및 건강 관련 특성 개선 연구 사례
밀 관련 질환과 관련된 글루텐 단백질이 밀 가공 품질을 조절하는 데 중요한 역할을 한다는 점을 감안하면 밀의 가공 품질과 건강 관련 특성을 동시에 개선하는 것은 밀 육종에 있어서 매우 중요하다. 밀의 글리아딘과 글루테닌의 구성 및 비율은 밀의 유전적 구성에 따라 달라지며, 현대 밀 육종 프로그램은 밀의 다양한 가공적성에 적합한 특성을 향상시키고 건강성을 개선하기 위해 글루텐 단백질 조성을 최적화하는 데 초점을 맞추고 있다(
Filip et al. 2023). 이러한 측면에서 글루텐 구성요소의 기능성을 개선하거나 위험성 글루텐 단백질을 제거 또는 변형함으로써 밀 품질 특성을 향상시키는 접근 방식을 검토하고 요약하였다.
물리 및 화학적 돌연변이원을 이용한 연구
돌연변이 유도(mutagenesis)는 돌연변이를 생성하여 생물체의 유전 정보를 변화시키는 효과적인 작물 개량 방식으로, 유전적 변이를 가속화하고 중요한 농업적 특성을 향상시킬 수 있다(
Pathirana 2011). 1950년대 이후에 전 세계적으로 밀 돌연변이 육종이 이루어졌고 현재까지 263종 이상의 밀 품종이 개발되었다(
Bakshi et al. 2020). 식물 돌연변이 유도를 위해서는 방사선 조사와 같은 물리적 돌연변이원(mutagen)과 알킬화제, 염기유사체, 탈아미노화제 등의 화학적 돌연변이원이 사용되고 있다. 여러 연구에서 밀의 면역원성 잠재력을 줄이거나 가공성을 개선하기 위해 돌연변이 유도 기술을 적용해왔다(
Tables 4,
5) (
Jouanin et al. 2019,
Kalia et al. 2000,
Kozub et al. 2013,
Lai et al. 2014,
Mangova & Rachovska 2004,
Nazarenko et al. 2023,
Rahemi et al. 2015,
Shaw et al. 2013,
Yang et al. 2023).
물리적 돌연변이원 중 가장 일반적으로 사용되는 것은 감마선(gamma ray, γ-ray)이며, 이는 밀 육종 프로그램에서 널리 사용된다. 감마선은 활성산소종(reactive oxygen species, ROS)을 유발하여 DNA에 산화 손상을 일으킴으로써 단일 또는 이중가닥 절단을 유도한다. 육배체 밀 품종 Paragon에 250 Gy 감마선을 조사한 계통에서 6A
Gli-2 유전자좌의 α-글리아딘과 1B의 γ-글리아딘에 대한 상동성 결실이 보고되었다(
Jouanin et al. 2019,
Shaw et al. 2013). 또한 건조된 밀 종자에 300 Gy의 선량으로 [
60Co] 감마선을 조사한 결과, 글루텐 함량과 HMW-GS 구성이 크게 변화하였으며, 이로 인해 밀가루의 반죽 혼합특성에 변화를 유도했다(
Lai et al. 2014). 일반 밀 종자에 200 Gy의 감마선을 조사한 연구에서는 글리아딘 유전자의 침묵이 발생하고 글리아딘에 대한 글루테닌의 비율이 증가하여 밀의 제빵 품질이 향상되었다(
Kozub et al. 2013). 감마선을 조사한 경우에도 제빵 특성이 향상되었으며, 대조군에 비해 단백질 함량, 습부량(wet gluten), 반죽 안정성(dough stability), 반죽 신장 저항성(resistance to dough extension)이 향상되었다(
Rahemi et al. 2015).
Kalia et al. (2000)은 아지리딘(aziridine), 니트로소화합물(nitro compound), 에스터(ester)를 포함한 다양한 화학적 돌연변이원을 사용하여 밀에서 돌연변이를 유도하고 그 효율에 대한 비교 분석을 수행했다. 그 결과 에틸메탄설포네이트(ethyl-methane sulfonate, EMS)가 밀의 펠스헨키값(Pelshenke value), 침전가(sedimentation value), 반죽 품질을 개선하는 효과적인 방법임을 확인했다. EMS는 주로 DNA에서 G/C에서 A/T 뉴클레오타이드로의 전이를 유도하는 화학적 돌연변이원으로 알려져 있다(
Jouanin et al. 2018). EMS 돌연변이를 통해 얻은 HMW-GS 결실 계통은 HMW-GS 함량이 현저히 감소하고 α-와 γ-글리아딘 함량이 증가했다(
Yang et al. 2023). 이에 따라 야생형과 비교했을 때 반죽 안정성 시간과 점탄성이 눈에 띄게 감소했다. 아지드화나트륨(sodium azide, 1 mM)을 사용하여 개량한 겨울 밀의 하이브리드-돌연변이 계통은 반죽 연화(dough softening), 안정성, 유동학적 품질이 더 높았으며(
Mangova & Rachovska 2004), 니트로소메틸요소(nitrosomethylurea, 0.0125%)를 적용하면 글리아딘과 글루테닌 함량이 증가하고 제빵 품질이 개선됨을 입증하였다(
Nazarenko et al. 2023).
RNA 간섭에 기반한 연구
RNA 간섭(RNA interference, RNAi)은 여러 생물체에서 보고된 서열 특이적 RNA 분해 시스템이며, 이중 가닥 RNA (double-stranded RNA, dsRNA)에 의한 전사 후 과정에서 비롯된 방어기작을 응용한 기술이다(
Gil-Humanes et al. 2008). RNA 간섭 기술은 글루텐 단백질 합성에 관여하는 특정 유전자를 침묵시키는 데 사용될 수 있는데, 주요 글루텐 단백질 유전자의 mRNA 전사체를 표적으로 삼아 분해하는 RNA 분자를 설계함으로써 특정 글루텐 단백질의 생산을 줄일 수 있다. 이 방식은 면역원성 감소를 목표로 하는 밀 품종 개발에 유용하다. 이전 연구들에 의하면 RNAi 기술을 적용한 밀 품종은 글리아딘 함량이 감소하고 면역원성이 낮으며 가공 품질이 개선되는 효과를 나타냈다(
Tables 4,
5) (
Altenbach & Allen 2011,
Altenbach et al. 2014a,
2014b,
2015,
2019,
2020b,
Barro et al. 2016,
Becher et al. 2012,
Blechl et al. 2016,
Gil-Humanes et al. 2008,
2010,
2012,
Pistón et al. 2011,
2013,
Sánchez-León et al. 2019,
Zhou et al. 2021). 미국 Altenbach 연구팀은 RNAi 기술을 사용하여 WDEIA와 관련된 주요 알레르기 유발 항원인 ω5-글리아딘을 감소시켰고 이러한 형질전환 밀의 잠재적 알레르기성을 평가했다(
Altenbach & Allen 2011,
Altenbach et al. 2015). 연구 결과에 따르면 ω5-글리아딘 유전자가 억제된 형질전환 밀 계통은 WDEIA 환자에서 IgE 반응이 감소했다. 한 계통에서는 ω1,2-글리아딘도 상당히 감소했을 뿐 아니라 다른 글루텐 단백질들도 약간 감소했으며, 다른 계통에서는 HMW-GS와 s-type LMW-GS가 감소했고 m-type LMW-GS와 α-글리아딘이 증가했다(
Altenbach et al. 2014b). 또한 형질전환 계통의 밀가루에서 반죽 혼합특성(dough mixing property)이 개선됨을 보여주었다(
Altenbach et al. 2014a). 빵밀(
Triticum aestivum cv. Butte 86)에서도 ω1,2-글리아딘 유전자의 5' 말단에 있는 141 bp 영역을 표적으로 하는 RNAi 플라스미드를 도입하여 유전자 변형이 이루어졌다(
Altenbach et al. 2019). 한 계통에서는 ω1,2-글리아딘이 생성되지 않았고 다른 단백질에는 영향을 미치지 않았다. 또 다른 계통에서는 거의 모든 글리아딘과 LMW-GS가 존재하지 않았지만, HMW-GS는 증가했으며 글로불린(globulin), 트리티신(triticin), 퓨리닌(purinin)과 같은 비글루텐 단백질이 증가하는 결과를 보였다. 셀리악병 환자 집단의 혈청 IgG (immunoglobulin G) 및 IgA (immunoglobulin A) 항체의 반응성은 두 형질전환 계통 모두에서 상당히 감소했고, 혼합 내성(mixing tolerance)은 ω1,2-글리아딘 결핍 계통에서 개선되었으나 글루텐 결핍이 가장 심한 계통에서는 감소했다.
γ-글리아딘이 억제된 RNAi-유도 형질전환 밀 계통에서도 γ-글리아딘은 감소했으나 아미노산 함량에는 영향을 미치지 않았고, 다른 글루텐 단백질이 증가하여 밀 반죽의 제빵 품질에 부정적인 영향을 미치지 않았음을 보여주었다(
Gil-Humanes et al. 2008,
Pistón et al. 2011). 동일 연구집단의 후속 연구에서 γ-글리아딘이 감소하면 다른 프롤라민의 생합성에 대한 보상 효과로 인해 반죽의 혼합 내성이 향상될 수 있음이 확인되었다(
Gil-Humanes et al. 2012).
Gil- Humanes et al. (2010)은 α-, ω-, γ-글리아딘 중에 고도로 보존된 서열을 포함하는 361 bp 키메라 단편으로 설계된 hairpin RNA (hpRNA)를 사용하여 빵밀 품종인 Bobwhite의 두 가지 유전자형에서도 형질전환 계통을 만들었다. 해당 연구는 키메라 단편이 세 그룹의 글리아딘 모두에서 유전자 발현을 효과적으로 하향 조절할 수 있음을 보여주었고, 모든 형질전환 계통에서 글리아딘 함량이 70-88% 범위로 상당히 감소했다. 모든 글리아딘을 침묵시킨 다른 연구에서는 글루테닌과 비글루텐 단백질이 증가하였지만, 총질소 함량에는 큰 영향을 미치지 않았다(
Pistón et al. 2013). 이어지는 후속 연구에서 글리아딘 함량이 낮은 계통의 빵은 일반 밀가루와 유사한 제빵 및 관능 특성을 가졌으나 최대 97%까지 글리아딘 함량이 낮아졌다(
Gil-Humanes et al. 2014a). 저글리아딘 밀가루는 또한 일반 밀가루보다 라이신(lysine) 함량이 현저히 높았으며, 주성분 분석(principal component analysis, PCA)을 통해 밀가루의 가공 특성을 평가한 결과, 전반적인 품질이 개선되었고 특히 반죽 안정성이 증가했다(
Gil-Humanes et al. 2014b).
Becker et al. (2012)은 α-글리아딘 서열의 5' 암호영역에서 유래한 313 bp의 DNA 단편을 hairpin-based RNAi 구성에 클로닝하여 형질전환을 수행하였으며, 그 결과 형질전환 계통은 α-글리아딘 수준 차이를 보였고, 변화가 없는 계통에서 심하게 감소한 계통까지 그 차이가 컸다. α-글리아딘 함량이 가장 크게 감소한 계통은 α-글리아딘의 결핍이 γ-와 ω-글리아딘, HMW-GS, 알부민/글로불린(albumins/globulins) 증가로 보상되었음을 보여주었다. 형질전환 계통은 글루텐 저항성(gluten resistance)이 현저히 증가하고 반죽의 신장성(extensibility)이 약간 감소했다. α-글리아딘과 HMW-GS 유전자를 침묵시킨 다른 연구에서는 모든 α-글리아딘이 효과적으로 침묵되었고 HMW-GS의 일부 오프타겟 유전자 침묵(off-target gene silencing)이 확인되었으며, 나머지 글루텐 단백질에 대한 보상 효과는 관찰되지 않았다(
Altenbach et al. 2020b).
Marín-Sanz et al. (2022)은 배유-특이적 프로모터에 의해 구동되는 글리아딘 유전자의 역반복 서열을 가진 플라스미드가 삽입된 야생형 BW208에서 RNAi 형질전환 밀 계통 D793과 D783을 파생시켰다. 두 품종은 NCGS와 관련된 글루텐과 프룩탄(fructan) 수치가 감소했을 뿐 아니라 비글루텐 단백질이 증가하고 스트레스 반응 기전에 차이가 있었다. RNAi 기술로 글리아딘 및 LMW-GS가 감소된 밀 계통에 대하여 셀리악병 환자의 말초혈액세포(peripheral blood mononuclear cell, PBMC)를 사용한 자극 반응도 조사되었다(
Sánchez-León et al. 2019). RNAi 형질전환 밀 계통 중에 특히 α- 및 γ-글리아딘과 DQ2.5 및 α-글리아딘 항원결정기 수준이 감소한 계통에서 면역원성이 감소하여 야생형 밀에 비해 PBMC 자극과 인터페론-감마(interferon-gamma) 방출이 낮아졌다.
α-, γ-, ω-글리아딘 및 LMW-GS의 RNAi 단편을 포함하는 7가지 플라스미드 조합이 프롤라민을 침묵시키는 효과에 대해서도 평가되었다(
Barro et al. 2016). 하향 조절된 글리아딘의 유형과 그 수준은 플라스미드 조합에 따라 달라졌고, 특히 세 가지 플라스미드 조합의 계통은 고도로 면역원성을 갖는 α-와 ω-글리아딘의 셀리악병 항원결정기가 전혀 없었다. 대부분 조합에서 HMW-GS는 상향 조절된 반면에 LMW-GS는 하향 조절되었으며, 사용된 플라스미드 조합과 관계없이 총단백질 및 전분 함량은 영향을 받지 않았다. 최근 연구에서 밀 글리아딘의 RNAi 침묵은 종자 저장단백질의 합성을 조절하는 전사인자 네트워크를 변화시켜 곡물 단백질 수준을 유지하는 것으로 밝혀졌다(
Marín-Sanz & Barro 2022). 글리아딘 단백질의 유전자 침묵으로 인해 LMW-GS가 감소하고 HMW-GS 수준은 변화가 없었으며, 글루텐이 아닌 단백질들의 함량이 증가하여 전체 질소 함량이 야생형과 비슷한 수준으로 유지되었다. RNAi를 통한 세칼린(secalin)의 하향 조절은 세칼린, ω-글리아딘, γ-글리아딘, LMW-GS, 파리닌(farinin)의 수준을 감소시켰고, α-글리아딘, 트리티신 및 y-type HMW-GS의 수준은 증가한 것으로 나타났다(
Blechl et al. 2016,
Zhou et al. 2021). 형질전환 계통의 밀가루는 반죽의 혼합특성에서 차이를 보였는데, 특히 반죽의 발달시간(dough development time)이 길어지고 혼합 내성이 더 커졌다. 이러한 연구 결과는 밀 글루텐 단백질 조절에 있어서 식물 생명공학과 유전자 변형 기술을 이해하는 데 중요한 통찰력을 제공하며, 글루텐 단백질 구성의 유전자 조작을 통해 건강성을 높이는 데 기여할 수 있다. 그러나 RNAi 기술은 그 특성상 목표 유전자 이외의 off-target 유전자에도 영향을 미칠 가능성이 높고, 이는 밀과 같은 복잡한 다배체(genome redundancy) 식물에서는 더욱 두드러질 수 있으며, 유사한 서열을 가진 다수의 글루텐 유전자들이 동시에 억제되는 multi-site suppression 현상이 발생할 수 있다(
Gil-Humanes et al. 2010,
McGinnis 2010,
Travella et al. 2006). 따라서 RNAi 기술은 특정 면역원성 펩타이드의 제거에는 효과적일 수 있으나, 동시에 비면역원성 글루텐 단백질의 발현도 억제하여 반죽 특성 및 제빵 품질에 영향을 미칠 수 있는 잠재적인 한계를 지닌다.
CRISPR/Cas9을 활용한 연구
크리스퍼(clustered regularly interspaced short palindromic repeats, CRISPR) 유전자가위는 생물체의 특정 DNA 서열을 인식하고 절단하여 정확하고 표적화된 변화를 가능하게 하는 혁신적인 유전체 편집 도구이다. 이 기술의 핵심 원리는 CRISPR와 Cas9 (CRISPR-associated protein 9) 단백질의 조합으로 이루어지며, 가이드 RNA (guide RNA, gRNA)를 사용하여 Cas9 효소를 특정 DNA 서열로 유도하여 절단하고 유전 물질을 삽입 또는 삭제한다(
Doudna & Charpentier 2014). 지금까지의 수많은 연구에 따르면 CRISPR/Cas9은 농업적 개량에서 의학적 차원에 이르기까지 다양한 목적으로 식물과 동물을 포함한 다양한 생물체의 유전자를 편집하는 데 효과적으로 사용되고 있다. CRISPR/Cas9 유전자 편집 시스템은 글리아딘 유전자에 의해 암호화되는 여러 항원결정기를 정확하게 수정함과 동시에 글리아딘 단백질의 가공 품질을 보존할 수 있는 잠재력이 있다.
밀에서 CRISPR/Cas9-매개 유전자 편집을 통해 글리아딘 축적에 주요 기여자로 확인된
Gli-γ1-1D와
Gli-γ2-1B 유전자를 녹아웃시킨 결과, 밀의 가공성이 향상되었고 셀리악병과 관련된 글루텐 항원결정기가 68-76%로 현저히 감소했다(
Table 4) (
Liu et al. 2023).
Sánchez-León et al. (2018)은 α-글리아딘 유전자를 표적으로 삼아 CRSPR/Cas9 유전자 편집 기술을 사용하여 저글루텐 형질전환 밀을 생성했으며, 돌연변이 계통의 α-글리아딘과 면역 반응성이 최대 85%까지 유의적으로 감소했고, 이러한 α-글리아딘의 현저한 감소가 반죽 점도를 향상시키는 데 기여하는 것으로도 밝혀졌다(
Table 5). 또 다른 연구에서는 특별히 설계된 단일 가이드 RNA (single guide RNA, sgRNA)의 CRISPR/Cas9를 사용하여 밀에서 α- 및 γ-글리아딘 유전자를 편집함으로써 글루텐 면역원성을 줄이는 것으로 목표했다(
Jouanin et al. 2019,
2020). 그들은 유전자 편집된 빵 밀 계통에서 글루텐 단백질의 명확한 변화를 관찰하였고, 글루텐이 감소했음에도 불구하고 편집된 밀 계통은 충분한 제빵 품질을 유지하여 셀리악병 환자를 위한 저면역원성 밀 품종을 개발할 수 있는 잠재력을 보여주었다. 최근 밀 품종 Fielder에서도 다중 CRISPR/Cas9 편집을 사용하여 글리아딘 유전자를 수정하였고, ω-와 γ-글리아딘 유전자 사본이 돌연변이 또는 삭제되고 α/β-글리아딘 유전자에서는 변화가 없었음을 보고했다(
Yu et al. 2023). 유전자 편집된 계통의 면역 반응성은 47배 감소했으며, 면역 반응성 모티프가 풍부한 ω-글리아딘(75%)과 γ-글리아딘(64%)이 유의적으로 감소하여 해당 밀의 면역독성 펩타이드 감소를 도모했다. 글리아딘이 감소했음에도 불구하고 편집된 계통의 가공 품질은 유지되었고 글루텐 중합 및 제빵 특성의 일부 매개변수도 개선되었다. 이 연구는 글루텐 유전자좌를 수정하는 강력한 도구로서 CRISPR-매개 유전자 편집의 유용성을 강조하여 밀의 면역 반응성을 줄이고 제빵 품질을 향상시킬 수 있는 잠재력을 제공한다.
글루텐 단백질 발현 조절과 관련 전사인자
밀에서 글루텐 단백질의 조절은 복잡한 과정으로 주로 여러 전사인자에 의해 전사 수준에서 조절된다. 이전 연구에서 밀에서 글루텐 단백질의 합성과 축적에 중요한 역할을 하는 몇 가지 주요 전사인자를 강조했다(
Makai et al. 2014,
Ravel et al. 2014). 글루텐 조절에 연관된 전사인자의 주목할만한 부분은 basic leucine zipper (bZIP)군이다. 이 단백질은 표적 유전자의 프로모터 영역에서 특정 DNA 서열에 결합한다. 예를 들어 storage protein activator (SPA)와 SPA heterodimerizing protein (SHP)은 글루테닌 유전자의 발현을 조절하는 두 가지 bZIP 유형 전사인자이다(
Éva et al. 2023,
Merlino et al. 2023). SPA는 글루테닌 유전자의 프로모터에서 GCN4-유사 모티프(GLM)와 G-box 모티프에 결합하여 전사를 활성화시키는 반면, SHP는 억제제 역할을 한다. 이러한 상호작용 특징은 밀 품질에 매우 중요한 글루테닌 유전자의 정확한 발현을 위한 복잡한 조절 네트워크를 이해하는 데 도움이 된다.
글루텐 조절에서 또 다른 중요한 전사인자 계열은 DNA-binding with one finger (DOF)군이다(
Mena et al. 1998). Wheat prolamin box binding factor (WPBF)는 이 계열의 주목할만한 구성원으로서 HMW-GS와 HMW-GS 유전자의 프로모터에 존재하는 prolamin-box (P-box) 또는 endosperm motif (EM) 서열에 결합하는 특징이 있다. WPBF는 밀에서 종자 저장단백질의 축적을 조절하는 데 중요한 역할을 하는데, SPA와 함께 종자 저장단백질 유전자의 전사를 가능하게 하여 글루텐 단백질의 합성을 촉진한다. 이에 더하여 scutellum and aleurone-expressed DOF (SAD) 전사인자도 최근 밀에서 글루텐 단백질 합성에 역할을 하는 것으로 입증되었다(
Merlino et al. 2023). 이는 글루테닌 유전자 프로모터의 특정 모티프에 결합하고 WPBF와 SPA를 포함한 다른 전사인자와 상호작용하여 유전자 발현을 조절하는 것으로 나타났다. 이러한 상호작용은 밀에서 글루텐 합성을 제어하는 복잡한 조절 메커니즘을 강조한다.
NAC 전사인자 계열도 글루텐 조절에 중요한 역할을 한다(
Gao et al. 2021). 이 계열에는 글루텐 단백질 조절을 포함한 다양한 발달 과정에 관여하는 식물 특정 전사인자가 포함된다. 여러 NAC 전사인자가 글루텐 단백질 합성에 참여한다는 사실이 기능적으로 밝혀졌다(
Merlino et al. 2023). 또한 R2R3MYB 계열 전사인자인 TaGAMYB는 프로모터의 특정 cis-조절 모티프에 결합하여 HMW-GS 유전자 발현을 직접 활성화하는 것으로 입증되었다. 이러한 다양한 전사인자의 공동 작용은 밀의 가공성에 필수적인 글루텐 단백질을 적절히 발현시키는 데 필수적이다. 이들 전사인자의 역할과 상호작용을 이해하는 노력을 통해 글루텐 조절의 유전적 및 분자적 기초에 대한 귀중한 통찰력을 얻을 수 있으며, 더 나아가 밀 육종 및 품질 개선 프로그램에 중요한 정보를 수집할 수 있다(
Liu et al. 2023). 그러나 글루텐 단백질의 발현은 프로모터와 같은 전사 조절 요인이나 외부 환경요인에 의해서도 영향을 받을 수 있기 때문에, 글루텐 면역원성 저감을 위한 접근은 단지 유전적 개량에 국한될 것이 아니라 여러 요인 변화에 따른 유전자 발현의 가변성까지 고려한 통합적인 전략이 요구된다(
Mikhaylenko et al. 2000,
Rogers et al. 2006).
비유전적 방법을 통한 글루텐 면역원성 저감
밀 글루텐 단백질의 면역원성 조절을 위한 접근법은 유전적 조작 외에도 비유전적인 방식으로 이루어질 수 있으며, 이는 소비자 수용성과 규제 측면에서 보다 실용적이고 적용 가능성이 높다. 대표적인 비유전적 방법으로는 효소 처리, 물리화학적 가공 기술, 발효 공정 등이 있다.
효소 처리는 글루텐 단백질의 구조적 안정성과 면역원성을 줄이기 위한 핵심기술 중 하나로, 다양한 단백질분해효소(protease)가 연구되고 있다(
Bradauskiene et al. 2021,
Caputo et al. 2010,
Tsiatsiani & Heck 2015). 펩신(pepsin), 트립신(trypsin), 키모트립신(chymotrypsin)과 같은 소화효소는 글루텐의 위장관 소화 과정을 모사하기 위해 사용되어 왔으며,
Caputo et al. (2010)은 특정 단백질분해효소를 활용하여 글리아딘 내 면역원성 항원결정기를 효과적으로 분해함으로써 셀리악병 유발 가능성을 낮출 수 있음을 제안하였다. 특히 최근 연구에서는 키위(
Actinidia deliciosa) 유래의 식물성 단백질분해효소인 액티니딘(actinidin)이 주목받고 있다.
Jayawardana et al. (2021,
2022)은 액티니딘이 인공적으로 합성된 33-mer 펩타이드와 프롤린 인접 결합을 포함한 펩타이드를 효과적으로 가수분해한다는 사실을 보고하였으며, 이는 기존의 글루텐 분해효소보다 높은 효율성을 보였다.
발효 공정 또한 글루텐 조절을 위한 효과적인 전략 중 하나이다. 특히 젖산균(lactic acid bacteria)과 같은 미생물은 발효 중 글루텐 펩타이드를 분해하거나 구조를 변화시켜 면역원성을 감소시킬 수 있다.
Di Cagno et al. (2004,
2010)은 특정 젖산균의 조합과 장시간 발효를 통해 밀가루 내 글루텐을 거의 완전히 분해하고 셀리악 환자에게 안전한 수준까지 면역반응 유발 가능성을 낮춘 발효 빵을 제조하는 데 성공했다. 이처럼 효소 처리와 다양한 가공 기술을 활용한 비유전적 전략은 밀 품종을 유전적으로 변형하지 않으면서도 글루텐 단백질의 구조 및 기능을 효과적으로 조절할 수 있는 실용적 수단으로 식품 산업 내 글루텐 민감 소비자층을 위한 대체 식품 개발에 있어 중요한 기술적 기반을 제공한다.
향후 전망 및 결론
글루텐은 밀을 토대로 한 생산물의 탄력성과 질감을 결정하는 데 중요한 역할을 하므로 제빵 품질에 절대적으로 중요하다. 그러나 셀리악병 및 WDEIA와 같은 글루텐 관련 질환이 증가함에 따라 밀의 글루텐 단백질을 줄이거나 그 구성을 수정하는 전략을 모색해야 할 필요성이 커지고 있다. 이를 통해 글루텐에 민감한 사람들에게 더 안전하면서도 기능성을 유지하는 밀 품종을 제공할 수 있다.
먼저, 돌연변이 육종은 엘리트 품종의 특정 형질을 표적하여 개량하면서도 기존 농업 형질 및 품질 특성을 보존할 수 있다(
Jouanin et al. 2018). 새로운 유전적 변이를 도입하고 육종 과정을 가속화하며 교배의 필요성을 없애 바람직하지 않은 특성이 도입될 위험을 줄일 수 있다. 더 나아가 분자표지이용선발(marker-assisted selection, MAS)과 유전체 편집과 같은 분자적 접근 방식과 함께 사용되어 식물 개량을 위한 정확하고 효율적인 방법을 제공할 수 있다. 그러나 유도된 돌연변이의 무작위성은 돌연변이 육종의 효능을 제한하여 유익한 결과 도출 빈도가 낮고, 의도하지 않은 특성이 발현될 가능성이 있어 시간 소모가 큰 과정이 될 수 있다. 게다가 유전자 편집의 정밀성이 부족하여 유전적 변화에 대한 제어 효율이 감소하고 유용한 돌연변이를 식별하기 위한 광범위한 스크리닝이 절대적으로 필요하다. 더욱이 상당한 수의 글리아딘 유전자와 그들의 밀접한 연관성으로 인하여 교배를 통해 단일 식물에 여러 글리아딘 유전자의 돌연변이를 도입하는 것은 매우 어렵다(
Jouanin et al. 2018). 결과적으로 돌연변이 육종 방식은 초기 계통에 이미 상당수의 돌연변이 된 글리아딘 유전자가 포함되어 있거나 많은 글리아딘이 제거된 경우에만 성공할 수 있다는 제한점이 있다.
RNAi 기술은 밀의 면역원성을 줄이는 데 큰 가능성을 보였지만, 결과적으로 생산되는 식물은 유전자가 변형되어 상업적 활용 가능성이 제한될 수 있다. 그에 반해 CRISPR/Cas9 기술은 외래 DNA 삽입 없이도 유전 가능한 돌연변이를 만들 수 있는 잠재력을 제공한다. 그럼에도 불구하고 CRISPR/Cas9을 통해 개발된 식물이 유전자 변형(genetically modified, GM) 작물로 분류될 것인지에 대한 문제는 여전히 논쟁의 여지가 있다. GM 작물에 대한 규제는 국가별로 상이하며 각국의 경제적, 환경적, 사회적 요인에 따라 차별적인 정책이 시행되고 있다(
Qaim 2016,
Turnbull et al. 2021). 미국과 캐나다, 브라질은 GM 작물 재배 및 식품 판매에 적극적인 반면, 한국과 일본, 유럽연합은 상업적인 GM 작물 재배를 금지하고 수입에 대해서도 엄격한 규제를 적용하고 있다(
Wohlers 2010,
2015). 중국과 인도는 GM 작물의 연구 및 기술 개발을 장려하면서 상업적 재배를 제한적으로 허용하고 있으며, 향후 GM 작물의 상업적 활용을 위한 정책 변화 가능성이 있다. 최근 GM 작물에 배타적이던 유럽과 아시아 일부 국가가 GM 작물에 더 유리한 규제 체계를 채택하기 시작했고, 유전자 편집의 지속적인 발전과 글루텐 관련 건강 문제에 대한 소비자의 인식이 높아짐에 따라 가까운 미래에 글루텐 면역원성이 낮은 밀이 육종 프로그램의 우선순위가 될 가능성이 커 보인다(
Park et al. 2019,
Turnbull et al. 2021). 그에 따라 건강 위험성을 낮추고 높은 가공 품질을 제공하는 유전자 편집 밀의 개량에 상당한 진전을 이룰 수 있을 것으로 사료된다.
CRISPR/Cas9을 유전체 편집에 적용하는 것은 여러 식물에서 검증되었지만, 밀에서 글루텐을 편집하기 위한 대규모 구현은 여전히 제한적이다. 주요 과제 중 하나는 밀 유전체의 복잡한 특성이다. 이질육배체 보통계밀(
T. aestivum)은 약 17 Gbp의 큰 유전체를 가지고 있고 반복적인 서열이 높은 비율로 차지하고 있다(
Verma et al. 2021). 이러한 반복적인 서열의 존재는 표적 돌연변이의 삽입을 방해하고 유전체 편집 과정을 더 복잡하게 만들기 때문에 상당한 과제로 남아있고, 더욱이 각 유전자는 세 개의 상동 사본(A, B, D 게놈)이 존재하기 때문에 유전자 편집 과정이 더 복잡해져 여러 유전자 사본을 동시에 표적화하는 것이 어려운 특징이 있다(
Sánchez-León et al. 2018). 또한 글루텐 관련 질환의 면역반응이 일부 주요 면역원성 펩타이드에 국한되지 않고, 다양한 글루텐 유래 항원결정기에 의해 유발될 수 있다(
Vader et al. 2002). 실제로 여러 연구에서 셀리악병 환자들이 α-, γ-, 및 ω-글리아딘 등 다양한 글루텐 단백질에 존재하는 다수의 항원결정기에 반응한다는 사실이 보고되었으며(
Alaedini & Green 2005,
kjbs-57-2-199-f et al. 2010), 환자 개별의 HLA-DQ2 또는 DQ8 유전형에 따라 면역반응의 중심이 되는 항원결정기가 달라질 수 있다(
Sollid 2002). 이러한 복합적인 면역반응 특성은 특정 유전자 편집 기술을 통해 제한된 수의 항원결정기만을 제거하는 접근이 충분하지 않을 수 있음을 시사한다. 따라서 면역원성을 효과적으로 완화하기 위해서는 표적 항원결정기 선정의 타당성, 항원결정기 간의 교차 반응성, 그리고 개인 간 면역반응의 이질성을 모두 고려한 종합적인 접근이 요구된다. 최근 복잡한 밀 유전체를 해독하는 데 있어 많은 진전이 이루어지면서 밀 품질을 개선할 수 있는 새로운 기회가 열렸으며(
IWGSC 2018,
Walkowiak et al. 2020), 유전체 편집 기술의 발전으로 복잡한 육배체 밀 유전체를 편집하기 위한 CRISPR의 잠재력이 높아졌다(
Camerlengo et al. 2020,
Jouanin et al. 2019,
Roychowdhury et al. 2024,
Wang et al. 2018,
Xie et al. 2015). 특정 코돈 최적화, 인트론 포함, 단백질 공학을 통해 밀에서 고효율의 동시 다중 유전자 편집을 가능하게 하는 최적화된 CRISPR 시스템이 확인되었고(
Lawrenson et al. 2024), 향상된 프라임 편집(prime editing) 기술은 밀의 편집 효율성을 크게 개선하여 식물에서 8개의 유전자를 동시 편집하여 여러 특성을 쌓을 수 있었다(
Ni et al. 2023). 유전자 편집 기술의 최근 발전은 글루텐 단백질 관련 건강 위험을 감소시키고 가공성이 향상된 밀 품종의 개발을 가속화하여 세계적인 식량안보를 강화하는 데 기여할 수 있을 것이라 기대한다. 또한 유전자 편집 작물과 GM 작물의 차이에 대한 소비자 이해도를 높이고 사회적 인식을 개선하며 안전성 검증이 지속적으로 이루어진다면, 향후 유전자 편집 밀 품종의 상용화와 시장 수용성이 더욱 확대될 것으로 전망된다.
적요
밀은 전 세계적으로 중요한 식량작물로서, 가공 다양성과 영양적 가치를 통해 식량안보에 크게 기여하고 있다. 그러나 글루텐 단백질은 셀리악병, 비셀리악 글루텐 민감성, 밀 알레르기 등과 같은 다양한 건강 문제와 관련이 있으며, 동시에 반죽의 점도나 탄력성과 같은 밀의 가공 특성에도 중요한 영향을 미치는 필수 요소이다. 현재 글루텐 관련 질환은 전 세계적으로 많은 사람에게 영향을 미치고 있기 때문에 높은 가공성을 유지하면서도 면역 반응성을 낮춘 밀 품종에 대한 요구를 증가시키고 있다. 본 논문은 글루텐 단백질 합성을 조절하는 분자적 기전과 글루텐 단백질의 화학 및 기능적 특성을 탐구하고, 글루텐 단백질을 변형하여 밀의 건강 및 품질 특성을 향상시키기 위한 최신 생명공학적 접근법의 진전을 요약한다. RNAi 및 CRISPR/Cas9과 같은 기술은 밀의 기능적 특성을 손상시키지 않으면서 면역 반응성을 줄이는 데 유망한 방법을 제공하며, 이러한 도구를 활용한 건강 향상과 고품질 밀이라는 이중 목표를 충족하는 밀 품종 개발에 대한 도전과제와 전망에 대해 논의한다.
사사
본 연구는 농촌진흥청 국립농업과학원의 기본연구사업(PJ01720903)의 지원으로 수행되었습니다.
Fig. 1Classification of gluten proteins by solubility and content of cysteine residue in common wheat. HMW-GS (high-molecular-weight glutenin subunit) and LMW-GS (low-molecular-weight glutenin subunit).
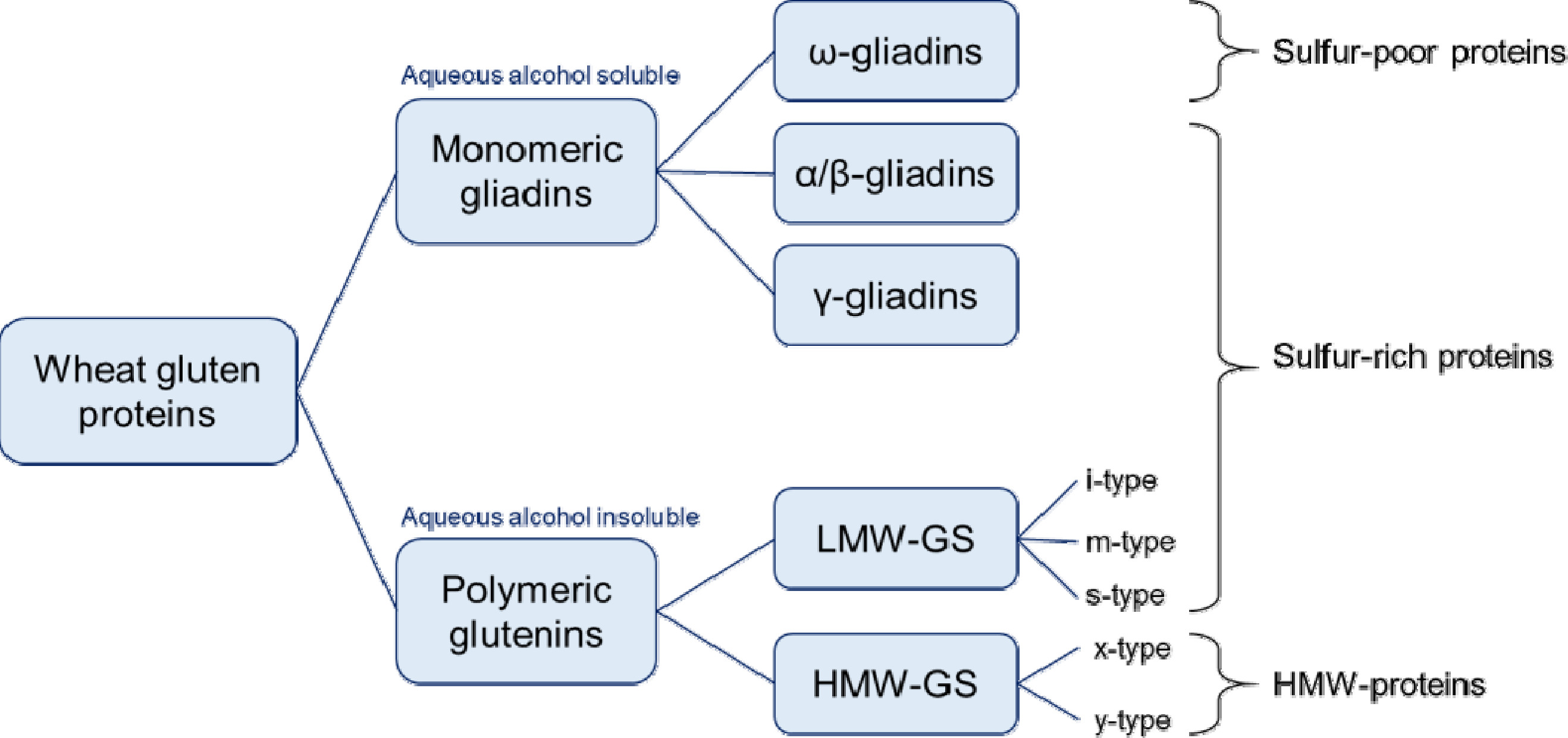
Fig. 2A representation of gluten protein structures as amino acids deduced from their genes. The position of cysteine residues involved in the formation of interchain and intrachain disulfide bonds is indicated by numbers. Cysteine residues forming intrachain disulfide bonds are linked in a chain.
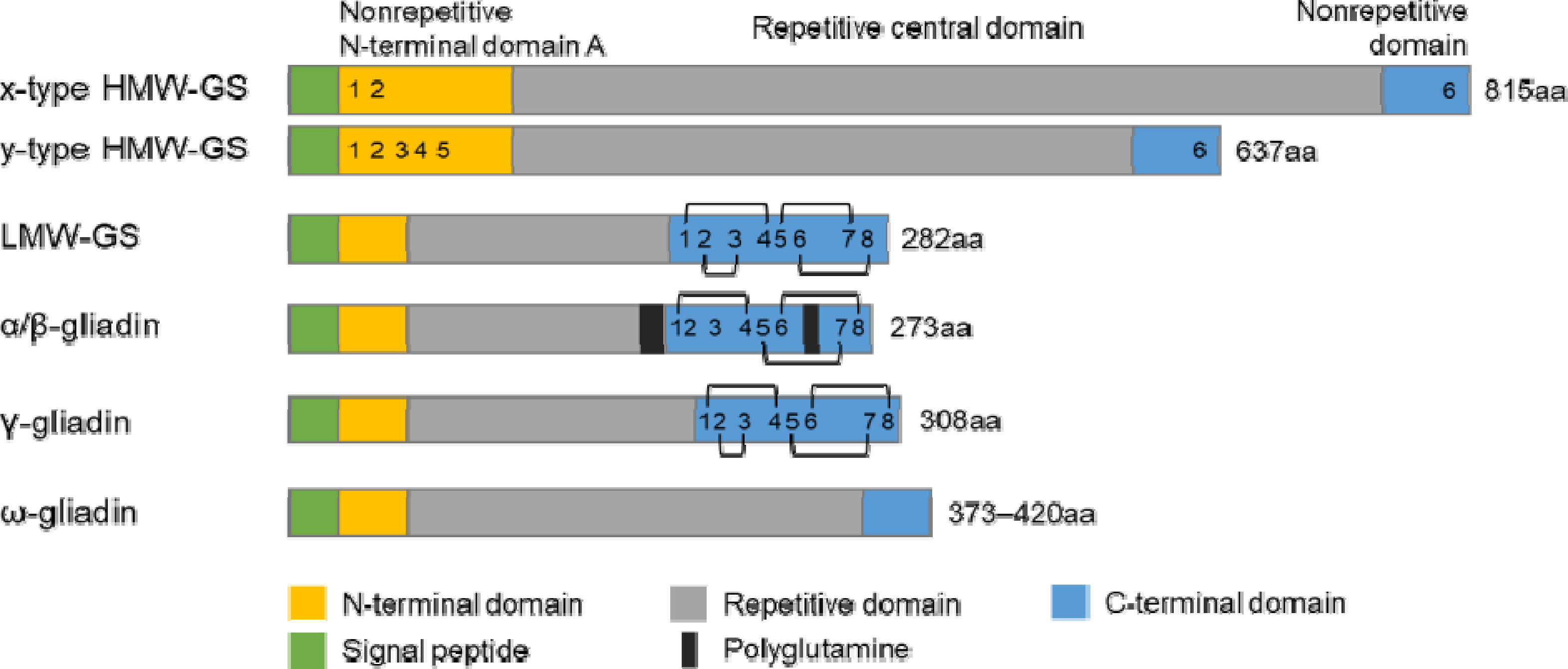
Fig. 3Diagrams representing the gluten polymers linked by forming interchain disulfide bonds between x- and y-type HMW-GS and LMW-GS.
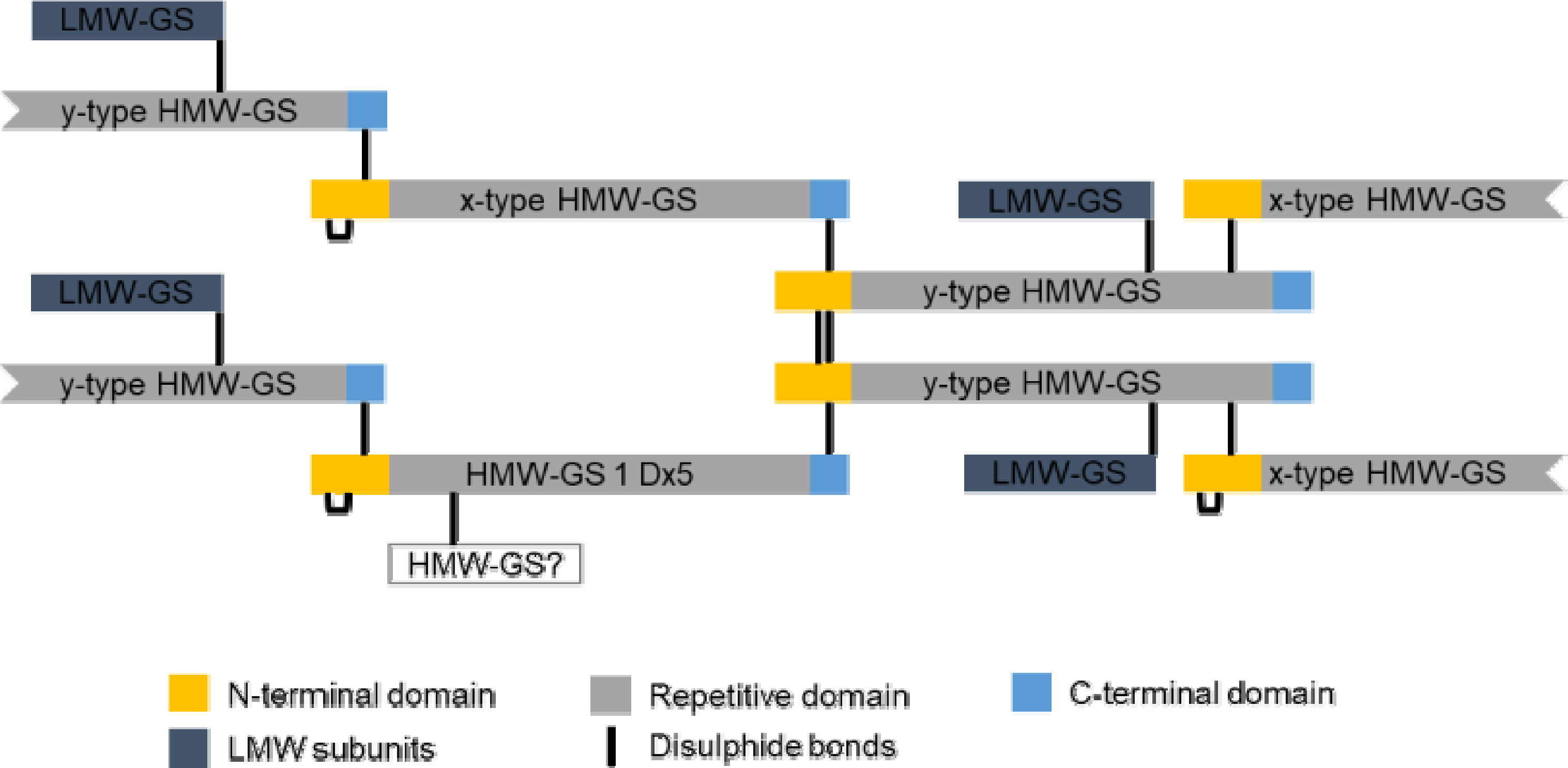
Fig. 4RP-UPLC profiles of gluten proteins in glutenin (A) and gliadin fraction (B) from reference wheat cultivar Chinese Spring.
Table 1Genetic loci of gluten proteins in common wheat.
Table 1
|
Protein type |
Chromosome arm |
Locus |
|
Gliadins |
|
|
|
γ-, ω5-, and ω1,2-gliadins |
1AS, 1BS, 1DS |
Gli-A1, Gli-B1, Gli-D1,
Gli-A3, Gli-B3, Gli-D3,
Gli-A2, Gli-B2, Gli-D2
|
|
α/β-gliadins |
6AS, 6BS, 6DS |
|
Glutenins |
|
|
|
HMW-GS |
1AL, 1BL, 1DL |
Glu-A1, Glu-B1, Glu-D1
|
|
LMW-GS |
1AS, 1BS, 1DS |
Glu-A3, Glu-B3, Glu-D3
|
Table 2Distribution of celiac disease (CD)-related epitopes in the full-length gluten genes of ‘Chinese Spring’.
Table 2
|
Gene |
Amino acid sequence of CD-relevant epitope |
The number of epitope |
|
α-gliadins |
|
|
|
α-A1 |
QGSFQPSQQ |
1 |
|
α-A2 |
QGSFQPSQQ |
1 |
|
α-A4 |
PFPQPQLPY, FRPQQPYPQ |
2 |
|
α-A5 |
PFPQPQLPY, FRPQQPYPQ |
2 |
|
α-A6 |
PFPQPQLPY, FRPQQPYPQ |
2 |
|
α-A8 |
PFPQPQLPY, FRPQQPYPQ |
2 |
|
α-A9 |
PFPQPQLPY |
1 |
|
α-A10 |
PFPQPQLPY, FRPQQPYPQ |
2 |
|
α-B3 |
QGSFQPSQQ |
1 |
|
α-B25 |
QGSFQPSQQ |
1 |
|
α-D1 |
QGSFQPSQQ |
1 |
|
α-D4 |
PFPQPQLPY, PQPQLPYPQ, FRPQQPYPQ |
3 |
|
α-D5z
|
PFPQPQLPY, PYPQPQLPY, PQPQLPYPQ (2), FRPQQPYPQ, QGSFQPSQQ |
6 |
|
α-D6 |
PFPQPQLPY, PYPQPQLPY, PQPQLPYPQ, QGSFQPSQQ |
4 |
|
α-D8 |
PFPQPQLPY, PYPQPQLPY, PQPQLPYPQ, FRPQQPYPQ, QGSFQPSQQ |
5 |
|
α-D9 |
PFPQPQLPY, PYPQPQLPY, PQPQLPYPQ, FRPQQPYPQ, QGSFQPSQQ |
5 |
|
γ-gliadins |
|
|
|
γ-A1 |
IQPQQPAQL, QQPQQPFPQ (6), QQPFPQQPQ (2), PFPQPQQPF |
10 |
|
γ-A3 |
PQQSFPQQQ, QQPQQPFPQ (4) |
5 |
|
γ-A4 |
PQQSFPQQQ, QQPQQPFPQ (4) |
5 |
|
γ-B1 |
PQQSFPQQQ, IQPQQPAQL, PQPQQQFPQ, QQPQQPFPQ (3), PQPQQPFCQ, PFPQPQQPF |
8 |
|
γ-B2 |
PQQSFPQQQ, IQPQQPAQL, PQPQQQFPQ, QQPQQPFPQ (3) |
6 |
|
γ-B4 |
PQQSFPQQQ, PQPQQQFPQ, QQPQQPFPQ (5), QQPFPQQPQ (3) |
10 |
|
γ-B6 |
PQQSFPQQQ, IQPQQPAQL, QQPQQPYPQ, QQPQQPFPQ (2) |
5 |
|
γ-D1 |
PQQSFPQQQ, IQPQQPAQL, QQPQQPFPQ (3), PQPQQPFCQ, PFPQPQQPF |
7 |
|
γ-D2 |
PQQSFPQQQ, IQPQQPAQL, QQPQQPFPQ (5), QQPFPQQPQ (3) |
10 |
|
γ-D3 |
PQQSFPQQQ, IQPQQPAQL, QQPQQPYPQ, SQPQQQFPQ, QQPQQPFPQ, QQPFPQQPQ |
6 |
|
γ-D4 |
PQQSFPQQQ, IQPQQPAQL, PQPQQQFPQ, QQPQQPFPQ (4), QQPFPQQPQ |
6 |
|
ω-gliadins |
|
|
|
ω-A4y
|
QQPQQPFPQ, QQPFPQQPQ (2) |
3 |
|
ω-D1 |
QQPQQPFPQ (5), QQPFPQQPQ (5), PFPQPQQPF, PQPQQPFPW |
12 |
|
ω-D2 |
QQPQQPFPQ (7), QQPFPQQPQ (7), PFPQPQQPF, PQPQQPFPW |
16 |
|
ω-D3 |
QQPQQPFPQ (7), QQPFPQQPQ (7), PFPQPQQPF, PQPQQPFPW |
16 |
|
LMW-GS |
|
|
|
LMW-A2 |
FSQQQQSPF |
1 |
|
LMW-B2 |
PFSQQQQPV |
1 |
|
LMW-B3 |
PFSQQQQPV (2) |
2 |
|
LMW-D1 |
PFSQQQQPV (2) |
2 |
|
LMW-D2 |
PFSQQQQPV |
1 |
|
LMW-D6 |
PFSQQQQPV (3) |
3 |
|
LMW-D7 |
PFSQQQQPV |
1 |
Table 3Distribution of WDEIA epitopes in the full-length gluten genes of ‘Chinese Spring’.
Table 3
|
Gene |
Amino acid sequence of WDEIA epitope |
The number of epitope |
|
ω-gliadins |
|
|
|
ω-B3 |
QQIPQQQ (3), QQFPQQQ (16), QQSPQQQ |
20 |
|
ω-B6 |
QQIPQQQ (4), QQFPQQQ (16), QQSPEQQ, QQSPQQQ |
22 |
|
ω-D4z
|
QQIPQQQ (5), QQFPQQQ (16) |
21 |
|
LMW-GS |
|
|
|
LMW-B3 |
QQFPQQQ |
1 |
|
LMW-B4 |
QQFPQQQ |
1 |
|
LMW-D6 |
QQFPQQQ |
1 |
Table 4Regulation of gluten proteins for reducing immunogenic potential in wheat.
Table 4
|
Method |
Mutagen |
Main findingz
|
Reference |
|
|
Mutagenesis |
Gamma-ray |
α- and γ-gliadins (-) |
Shaw et al. 2013
|
|
|
Method |
Target gene |
Main finding |
Reference |
|
RNAi |
ω5-gliadins |
ω5- and ω1,2-gliadins (-) |
Altenbach et al. 2014a
|
|
ω5-gliadins |
ω5- and ω1,2-gliadins, HMW-GS, s-type LMW-GS (-);α-gliadins, m-type LMW-GS (+) |
Altenbach et al. 2014b
|
|
ω5-gliadins |
ω5-gliadins, IgE responses (-) |
Altenbach & Allen 2011, Altenbach et al. 2015
|
|
ω1,2-gliadins |
LMW-GS (-);HMW-GS, non-gluten proteins (globulins, triticins, purinins) (+) |
Altenbach et al. 2019
|
|
γ-gliadins |
γ-gliadins (-); other gluten proteins (+); amino acid (0) |
Gil-Humanes et al. 2008, Pistón et al. 2011
|
|
α-gliadins |
α-gliadins, dough extensibility (-);ω- and γ-gliadins, HMW-GS, albumins/globulins, gluten resistance (+) |
Becker et al. 2012
|
|
α-, ω-, and γ-gliadins |
α-, ω-, and γ-gliadins (-); HMW-GS (+) |
Gil-Humanes et al. 2010, 2014a, b, Pistón et al. 2013 |
|
α/β-, ω-, and γ-gliadins |
Gliadins, fructans linked with NCGS (-) |
Marín-Sanz et al. 2022
|
|
α-, ω-, and γ-gliadins |
Gliadins, LMW-GS (-);non-gluten proteins, total nitrogen (+); HMW-GS (0) |
Marín-Sanz & Barro. 2022
|
|
α-gliadins, HMW-GS |
α-gliadins, HMW-GS, reactivity of IgG and IgA antibidies, mixing properties, dough strength (-) |
Altenbach et al. 2020b
|
|
α-, ω-, and γ-gliadins, LMW-GS |
α-, ω-, and γ-gliadins, LMW-GS (-); HMW-GS (+);total protein, starch contents (0) |
Barro et al. 2016
|
|
α/β-, ω-, and γ-gliadins, LMW-GS |
Gliadins, LMW-GS, key epitopes (DQ2.5) (-); HMW-GS (+) |
Sánchez-León et al. 2019
|
|
Secalins |
Secalins, ω- and γ-gliadins, LMW-GS, farinins (-);α-gliadins, triticins, y-type HMW-GS (+) |
Blechl et al. 2016
|
|
ω-secalins |
ω-secalins, ω- and γ-gliadins (-); α-gliadins, HMW-GS (±) |
Zhou et al. 2021
|
|
CRISPR/Cas9 |
Gli-γ1-1D, Gli-γ2-1B
|
γ-gliadins, gluten epitopes (-) |
Liu et al. 2023
|
|
α-gliadins |
α-gliadins, immunoreactivity (-) |
Sánchez-León et al. 2019
|
|
a- and γ-gliadins |
α-gliadins, immunogenicity (-); baking quality (0) |
Jouanin et al. 2019, 2020
|
|
ω- and γ-gliadins |
ω- and γ-gliadins, immunoreactivity (-);α/β-gliadins, end-use quality (0) |
Yu et al. 2023
|
Table 5Regulation of gluten proteins for improving wheat quality related to end-use traits.
Table 5
|
Method |
Mutagen |
Main findingz
|
Reference |
|
Mutagenesis |
Gamma-ray |
Gliadins, ratio of glutenin to gliadin, baking quality (+) |
Kozub et al. 2013
|
|
Gamma-ray |
Wet gluten, dough stability, resistance to dough extension (+) |
Rahemi et al. 2015
|
|
Gamma-ray |
Wet gluten, HMW-GS, Farinograph parameters (±) |
Liu et al. 2023
|
|
EMS |
HMW-GS, dough stability time, viscoelasticity (-);α- and γ-gliadins (+) |
Yang et al. 2023
|
|
EMS |
Pelshenke value, sedimentation value, dough quality (+) |
Kalia et al. 2000
|
|
Nitrosomethylureas |
Gliadins, glutenins, baking quality (+) |
Nazarenko et al. 2023
|
|
Sodium azide |
Dough softening, dough stability, rheological quality (+) |
Mangova & Rachovska 2004
|
|
|
Method |
Target gene |
Main finding |
Reference |
|
|
RNAi |
ω5-gliadins |
ω5- and ω1,2-gliadins (-); mixing time and tolerance (+) |
Altenbach et al. 2014a
|
|
γ-gliadins |
Mixing tolerance, other prolamines (+) |
Gil-Humanes et al. 2012
|
|
α-, ω-, and γ-gliadins |
α-, ω-, and γ-gliadins (-);HMW-GS, lysine, dough and nutritional properties (+);total nitrogen, baking and organoleptic properties (0) |
Gil-Humanes et al. 2010, 2014a, b, Pistón et al. 2013
|
|
Secalins |
Secalins, y-type HMW-GS, mixing properties (+) |
Blechl et al. 2016
|
|
CRISPR/Cas9 |
Gli-γ1-1D, Gli-γ2-1B
|
γ-gliadins (-); end-use quality (+) |
Liu et al. 2023
|
|
α-gliadins |
Dough viscosity (+) |
Sánchez-León et al. 2019
|
References
- 1. Alaedini A, Green PHR. 2005. Narrative review: celiac disease: Understanding a complex autoimmune disorder. Ann Intern Med 142: 289-298.
- 2. Altenbach SB, Allen PV. 2011. Transformation of the US bread wheat 'Butte 86' and silencing of omega-5 gliadin genes. GM Crops 2: 66-73.
- 3. Altenbach SB, Chang HC, Rowe MH, Yu XB, Simon-Buss A, Seabourn BW, Green PH, Alaedini A. 2020b. Reducing the immunogenic potential of wheat flour: silencing of alpha gliadin genes in a U.S. wheat cultivar. Front Plant Sci 11: 20
- 4. Altenbach SB, Chang HC, Simon-Buss A, Mohr T, Huo N, Gu YQ. 2020a. Exploiting the reference genome sequence of hexaploid wheat: A proteomic study of flour proteins from the cultivar Chinese Spring. Funct Integr Genomics 20: 1-16.
- 5. Altenbach SB, Chang HC, Siomon-Buss A, Jang YR, Denery-Papini S, Pineau F, Gu YQ, Huo N, Lim SH, Kang CS, Lee JY. 2018. Towards reducing the immunogenic potential of wheat flour: omega gliadins encoded by the D genome of hexaploidy wheat may also harbor epitopes for the serious food allergy WDEIA. BMC Plant Biol 18: 291
- 6. Altenbach SB, Chang HC, Yu XB, Seabourn BW, Green PH, Alaedini A. 2019. Elimination of omega-1,2 gliadins from bread wheat (Triticum aestivum) flour: Effects on immunogenic potential and end-use quality. Front Plant Sci 10: 580
- 7. Altenbach SB, Tanaka CK, Allen PV. 2014b. Quantitative proteomic analysis of wheat grain proteins reveals differential effects of silencing of omega-5 gliadin genes in transgenic lines. J Cereal Sci 59: 118-125.
- 8. Altenbach SB, Tanaka CK, Pineau F, Lupi R, Drouet M, Beaudouin E, Morisset M, Denery-Papini S. 2015. Assessment of the allergenic potential of transgenic wheat (Triticum aestivum) with reduced levels of ω5-gliadins, the major sensitizing allergen in wheat-dependent exercise-induced anaphylaxis. J Agric Food Chem 63: 9323-9332.
- 9. Altenbach SB, Tanaka CK, Seabourn BW. 2014a. Silencing of omega-5 gliadins in transgenic wheat eliminates a major source of environmental variability and improves dough mixing properties of flour. BMC Plant Biol 14: 393
- 10. Altpeter F, Vasil V, Srivastava V, Vasil IK. 1996. Integration and expression of the high-molecular-weight glutenin subunit 1Ax1 gene into wheat. Nat Biotechnol 14: 1155-1159.
- 11. Anderson OD, Greene FC. 1989. The characterization and comparative analysis of high-molecular-weight glutenin genes from genomes A and B of a hexaploid bread wheat. Theor Appl Genet 77: 689-700.
- 12. Anderson OD, Gu YQ, Kong X, Lazo GR, Wu J. 2009. The wheat ω-gliadin genes: Structure and EST analysis. Funct Integr Genomics 9: 397-410.
- 13. Anjum FM, Khan MR, Din A, Saeed M, Pasha I, Arshad MU. 2007. Wheat gluten: high molecular weight glutenin subunits-structure, genetics, and relation to dough elasticity. J Food Sci 72: 56-63.
- 14. Avni R, Nave M, Barad O, Baruch K, Twardziok SO, Gundlach H, Hale I, Mascher M, Spannagl M, Wiebe K, Jordan KW, Golan G, Deek J, Ben-zvi B, Ben-zvi G, Himmelbach A, Maclachlan RP, Sharpe AG, Fritz A, Ben-david R, Budat H, Fahima T, Korol A, Faris JD, Hernandez A, Mikel MA, Levy AA, Steffenson B, Maccaferri M, Tuberosa R, Cattivelli L, Faccioli P, Ceriotti A, Kashkush K, Pourkheirandish M, Komatsuda T, Eilam T, Sela H, Sharon A, Ohad N, Chamovitz DA, Mayer KFX, Stein N, Ronen G, Peleg Z, Pozniak CJ, Akhunov ED, Distelfeld A. 2017. Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 357: 93-97.
- 15. Bakshi S, Jambhulkar SJ, Kumar U, Bhati P. In: Sareen S, Sharma P, Singh C, Jasrotia P, Singh GP, Sarial AK. 2020. (Eds). Induced mutagenesis to sustain wheat production under changing climate. . Improving cereal productivity through climate smart practices. Elsevier, Amsterdam, Netherlands. pp. s 37-63,
- 16. Barak S, Mudgil D, Khatkar BS. 2015. Biochemical and functional properties of wheat gliadins: A review. Crit Rev Food Sci Nutr 55: 357-368.
- 17. Barro F, Iehisa JCM, Giménez MJ, García-Molina MD, Ozuna CV, Comino I, Sousa C, Gil-Humanes J. 2016. Targeting of prolamins by RNAi in bread wheat: Effectiveness of seven silencing-fragment combinations for obtaining lines devoid of coeliac disease epitopes from highly immunogenic gliadins. Plant Biotechnol J 14: 986-996.
- 18. Battais F, Mothes T, Moneret-Vautrin DA, Pineau F, Kanny G, Popineau Y, Bodinier M, Denery-Papini S. 2005. Identification of IgE-binding epitopes on gliadins for patients with food allergy to wheat. Allergy 60: 815-821.
- 19. Becker D, Wieser H, Koehler P, Folck A, Mühling KH, Zörb C. 2012. Protein composition and techno-functional properties of transgenic wheat with reduced α-gliadin content obtained by RNA interference. J Appl Bot Food Qual 85: 23-33.
- 20.
- 21. Biesiekierski JR, Muir JG, Gibson PR. 2013. Is gluten a cause of gastrointestinal symptoms in people without celiac disease? Curr Allergy Asthma Rep 13: 631-638.
- 22. Blechl A, Beecher B, Vensel W, Tanaka C, Altenbach S. 2016. RNA interference targeting rye secalins alters flour protein composition in a wheat variety carrying a 1BL⋅1RS translocation. J Cereal Sci 68: 172-180.
- 23. Bradauskiene V, Vaiciulyte-Funk L, Shah BR, Cernauskas D, Tita MA. 2021. Recent advances in biotechnological methods for wheat gluten immunotoxicity abolishmenet-a review. Pol J Food Nutr Sci 71: 5-20.
- 24. Branlard G, Dardevet M, Saccomano R, Lagoutte F, Gourdon J. 2001. Genetic diversity of wheat storage proteins and bread wheat quality. Euphytica 119: 59-67.
- 25. Brzozowski B, Stasiewicz K, Adamczak M. 2023. Biotransformation of wheat proteins with Lactobacillus sp. following high hydrostatic pressure pre-treatment to reduce gluten immunoreactivity. J Cereal Sci 114: 103787.
- 26. Butow BJ, Ma W, Gale KR, Cornish GB, Rampling L, Larroque O, Morell MK, Békés F. 2003. Molecular discrimination of Bx7 alleles demonstrates that a highly expressed high-molecular-weight glutenin allele has a major impact on wheat flour dough strength. Theor Appl Genet 107: 1524-1532.
- 27. Camerlengo F, Frittelli A, Sparks C, Doherty A, Martignago D, Larré C, Lupi R, Sestili F, Masci S. 2020. CRISPR-Cas9 multiplex editing of the α-amylase/trypsion inhibitor genes to reduce allergen proteins in durum wheat. Front Sustain Food Syst 4: 104
- 28. Caputo I, Lepretti M, Martucciello S, Esposito C. 2010. Enzymatic strategies to detoxify gluten: Implications for celiac disease. Enzyme Res 2010: 174354.
- 29. Cha RR, Kim HJ. 2020. Non-celiac gluten sensitivity. Korean J Gastroenterol 75: 11-16.
- 30. Chlubnová M, Christophersen AO, Sandve GKF, Lundin KEA, Jahnsen J, Dahal-Koirala S, Sollid LM. 2023. Identification of gluten T cell epitopes driving celiac disease. Sci Adv 9: eade5800.
- 31. Cornish GB, Békés F, Eagles HA, Payne PI. In: Wrigley C, Békés F, Bushuk W. 2006. (Eds). Prediction of dough properties for bread wheats. . Gliadin and glutenin: The unique balance of wheat quality. AACC International Press, MN, USA. pp. 243-280,
- 32. Cummins AG, Roberts-Thomson ICR. 2009. Prevalence of celiac disease in the Asia-Pacific region. J Gastroenterol Hepatol 24: 1347-1351.
- 33. D′Ovidio R, Masci S. 2004. The low-molecular-weight glutenin subunits of wheat gluten. J Cereal Sci 39: 321-339.
- 34. Delcour JA, Joye IJ, Pareyt B, Wilderjans E, Brijs K, Lagrain B. 2012. Wheat gluten functionality as a quality determinant in cereal-based food products. Annu Rev Food Sci Technol 3: 469-492.
- 35. Di Cagno R, Barbato M, Di Camillo C, Rizzello CG, Angelis MD, Giuliani G, De Vincenzi M, Gobbetti M, Cucchiara S. 2010. Gluten-free sourdough wheat baked goods appear safe for young celiac patients: A pilot study. J Pediatr Gastroenterol Nutr 51: 777-783.
- 36. Di Cagno R, De Angelis M, Auricchio S, Greco L, Clarke C, De Vincenzi M, Giovannini C, D′Archivio M, Landolfo F, Parrilli G, Minervini F, Arendt E, Gobbetti M. 2004. Sourdough bread made from wheat and nontoxic flours and started with selected lactobacilli is tolerated in celiac sprue patients. Appl Environ Microbiol 70: 1088-1096.
- 37. Don C, Mann G, Bekes F, Hamer RJ. 2006. HMW-GS affect the properties of glutenin particles in GMP and thus flour quality. J Cereal Sci 44: 127-136.
- 38. Dong L, Zhang X, Liu D, Fan H, Sun J, Zhang Z, Qin H, Li B, Hao S, Li Z, Wang D, Zhang A, Ling HQ. 2010. New insights into the organization, recombination, expression and functional mechanism of low molecular weight glutenin subunit genes in bread wheat. PLOS ONE. 5: e13548.
- 39. Dong Z, Yang Y, Li Y, Zhang K, Lou H, An X, Dong L, Gu YQ, Anderson OD, Liu X, Qin H, Wang D. 2013. Haplotype variation of Glu-D1 locus and the origin of Glu-D1d allele conferring superior end-use qualities in common wheat. PLOS ONE 8: e74859.
- 40. Dong Z, Yang Y, Zhang K, Li Y, Wang J, Wang Z, Liu X, Qin H, Wang D. 2017. Development of a new set of molecular markers for examining Glu-A1 variants in common wheat and ancestral species. PLOS ONE 12: e0180766.
- 41. Doudna JA, Charpentier E. 2014. The new frontier of genome engineering with CRISPR-Cas9. Science 346: 1258096.
- 42. Éva C, Moncsek B, Szőke-Pázsi K, Kunos V, Mészáros K, Sági L, Juhász A. Makai2023. bZIP transcription factors repress the expression of wheat (Triticum aestivum L.) high molecular weight glutenin subunit genes in vegetative tissues. Acta Physiol Plant 45: 29
- 43. Faihs V, Kugler C, Schmalhofer V, Scherf KA, Lexhaller B, Mortz CG, Bindslev-Jensen C, Biedermann T, Brockow K. 2023. Wheat-dependent exercise-induced anaphylaxis: Subtypes, diagnosis, and management. J Dtsch Dermatol Ges 21: 1131-1135.
- 44. FAOSTAT.2022. Food and agriculture organization of the united nations.
- 45. Farahani H, Moghaddam ME, Mohammadi A, Nezhad KZ, Naghipour F. 2023. Effect of high molecular weight glutenin subunits on wheat quality properties, across a wide range of climates and environments in Iran. Physiol Mol Biol Plants 29: 889-902.
- 46. Ferranti P, Mamone G, Picariello G, Addeo F. 2007. Mass spectrometry analysis of gliadins in celiac disease. J Mass Spectrom 42: 1531-1548.
- 47. Filip E, Woronko K, Stępień E, Czarniecka N. 2023. An overview of factors affecting the functional quality of common wheat (Triticum aestivum L.). Int J Mol Sci 24: 7524
- 48. Ganbaatar U, Bodisad G, Jambal T, Noov B, Dorjgotov D, Subedi M. 2025. Allelic variation in HMW-GS of mongolian wheat varieties using KASP molecular marker. Plant Breed Biotech 13: 5-14.
- 49. Gao X, Liu T, Ding M, Wang J, Li C, Wang Z, Li X. 2018. Effects of HMW-GS Ax1 or Dx2 absence on the glutenin polymerization and gluten micro structure of wheat (Triticum aestivum L.). Food Chem 240: 626-633.
- 50. Gao Y, An K, Guo W, Chen Y, Zhang R, Zhang X, Chang S, Rossi V, Jin F, Cao X, Xin M, Peng H, Hu Z, Guo W, Du J, Ni Z, Sun Q, Yao Y. 2021. The endosperm-specific transcription factor TaNAC019 regulates glutenin and starch accumulation and its elite allele improves wheat grain quality. Plant Cell 33: 603-622.
- 51.
- 52. Gil-Humanes J, Pistón F, Barro F, Rosell CM. 2014b. The shutdown of celiac disease-related gliadin epitopes in bread wheat by RNAi provides flours with increased stability and better tolerance to over-mixing. PLOS ONE 9: e91931.
- 53. Gil-Humanes J, Pistón F, Giménez MJ, Martín A, Barro F. 2012. The introgression of RNAi silencing of γ-gliadins into commercial lines of bread wheat changes the mixing and technological properties of the dough. PLOS ONE 7: e45937.
- 54. Gil-Humanes J, Pistón F, Hernando A, Alvarez JB, Shewry PR, Barro F. 2008. Silencing of γ-gliadins by RNA interference (RNAi) in bread wheat. J Cereal Sci 48: 565-568.
- 55. Gil-Humanes J, Pistòn F, Tollefsen S, Sollid LM, Barro F. 2010. Effective shutdown in the expression of celiac disease-related wheat gliadin T-cell epitopes by RNA interference. Proc Natl Acad Sci 107: 17023-17028.
- 56. Gu YQ, Salse J, Coleman-Derr D, Dupin A, Crossman C, Lazo GR, Huo N, Belcram H, Ravel C, Charmet G, Charles M, Anderson OD, Chalhoub B. 2006. Types and rates of sequence evolution at the high-molecular-weight glutenin locus in hexaploid wheat and its ancestral genomes. Genetics 174: 1493-1504.
- 57. Gupta PK, Mir RR, Mohan A, Kumar J. 2008. Wheat genomics: Present status and future prospects. Int J Plant Genom 2008: 896451.
- 58. Huo N, Zhang S, Zhu T, Dong L, Mohr T, Hu T, Liu Z, Dvorak J, Luo MC, Wang D, Lee JY, Altenbach S, Gu YQ. 2018. Gene duplication and evolution dynamics in the homeologous regions harboring multiple prolamin and resistance gene families in hexaploid wheat. Front Plant Sci 9: 673
- 59. Huo Z, Zhu T, Altenbach S, Dong L, Wang Y, Mohr T, Liu Z, Dvorak J, Luo MC, Gu YQ. 2018. Dynamic evolution of α-gliadin prolamin gene family in homeologous genomes of hexaploid wheat. Sci Rep 8: 5181
- 60. Ikeda TM, Yanaka M, Takata K. 2014. Comparison of quality-related alleles among Australian and North American wheat classes exported to Japan. Cereal Chem 91: 616-622.
- 61. International Wheat Genome Sequencing Consortium (IWGSC)2018. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 361: eaar7191.
- 62. Jayawardana IA, Boland MJ, Higgs K, Zou M, Loo T, McNabb WC, Montoya CA. 2021. The kiwifruit enzyme actinidin enhances the hydrolysis of gluten proteins during simulated gastrointestinal digestion. Food Chem 341: 128239.
- 63. Jayawardana IA, Boland MJ, Loo TS, McNabb WC, Montoya CA. 2022. Actinidin reduces gluten-derived immunogenic peptides reaching the small intestine in an in vitro semi-dynamic gastrointestinal tract digestion model. Food Res Int 159: 111560.
- 64. Jeong KG, Kim JH, Ahn KM, Lee SY, Min TK, Pyun BY, Kim YH, Kim KW, Sohn MH, Kim KE, Jang GC, Song TW, Kim JH, Jeon YH, Lee YJ, Par YM, Yum HY, Kim HH, Kim WK, Lee SY. 2017. Age-based causes and clinical characteristics of immediate-type food allergy in Korean children. Allergy Asthma Immunol Res 9: 423-430.
- 65. Jouanin A, Gilissen LJWJ, Boyd LA, Cockram J, Leigh FJ, Wallington EJ, van den Broeck HC, van der Meer IM, Schaart JG, Visser RGF, Smulders MJM. 2018. Food processing and breeding strategies for coeliac-safe and healthy wheat products. Food Res Int 110: 11-21.
- 66. Jouanin A, Gilissen LJWJ, Schaart JG, Leigh FJ, Cockram J, Wallington EJ, Boyd LA, van den Broeck HC, van der Meer IM, America AHP, Visser RGF, Smulders MJM. 2020. CRISPR/Cas9 gene editing of gluten in wheat to reduce gluten content and exposure-reviewing methods to screen for coeliac safety. Front Plant Sci 7: 51
- 67. Jouanin A, Schaart JG, Boyd LA, Cockram J, Leigh FJ, Bates R, Wallington EJ, Visser RGF, Smulders MJM. 2019. Outlook for coeliac disease patients: towards bread wheat with hypoimmunogenic gluten by gene editing of α- and γ-gliadin gene families. BMC Plant Biol 19: 333
- 68. Kalia CS, Kharkwal MC, Singh MP. 2000. Recovery of desirable mutations in wheat. Indian J Genet 60: 465-470.
- 69. Karaduman Y, Yeşildağ ZS, Akın A. 2022. Evaluating selection efficacy of high molecular weight glutenin subunits (HMWGs) by relating gluten quality parameters. LWT 155: 112949.
- 70. Khan H. In: Qureshi A, Dar Z, Wani S. 2019. (Eds). Genetic improvement for end-use quality in wheat. . Quality breeding in field crops. Springer, Cham. pp. 239-253,
- 71. Köhler P, Belitz HD, Wieser H. 1991. Disulphide bonds in wheat gluten: isolation of a cystine peptide from glutenin. Z Lebensm Unters Forch 192: 234-239.
- 72. Köhler P, Belitz HD, Wieser H. 1993. Disulphide bonds in wheat gluten: further cystine peptides from high molecular weight (HMW) and low molecular weight (LMW) subunits of glutenin and from gamma-gliadins. Z Lebensm Unters Forch 196: 239-247.
- 73. Koike Y, Yanagida N, Sato S, Asaumi T, Ogura K, Ohtani K, Imai T, Ebisawa M. 2018. Predictors of persistent wheat allergy in children: a retrospective cohort study. Int Arch Allergy Immunol 176: 249-254.
- 74. Kozub NA, Sozinov IA, Blume YB, Sozinov AA. 2013. Study of the effects produced by gamma-irradiation of common wheat F1 seeds using gliadins as genetic markers. Cytol Genet 47: 13-19.
- 75. KREI2024. World Grain Market. 13: 5-29.
- 76. Lai DE, Wang M, Zhang CY. 2014. Quality trait variations in [60Co]-irradiated wheat and high-molecular-weight glutenin subunit mutant identification. Genet Mol Res 13: 9024-9031.
- 77. Lawrenson T, Clarke M, Kirby R, Forner M, Steuernagel B, Harwood W. 2024. An optimised CRISPR Cas9 and Cas12a toolkit for barley and wheat. Plant Methods in press.,
- 78. Lee HS, Lee KJ. 2019. Diagnosis of food hypersensitivity and food intolerance in patients with a functional gastrointestinal disorder. Korean J Med 94: 438-442.
- 79. Lee JY, Kim YT, Kang CS, Lim SH, Ha SH, Ahn SN, Kim YM. 2011. High molecular weight glutenin subunit in common wheat (Triticum aestivum L.). Korean J Breed Sci 43: 479-489.
- 80. Li Y, Fu J, Shen Q, Yang D. 2021. High-molecular-weight glutenin subunits: genetics, structures, and relation to end use qualities. Int J Mol Sci 22: 184
- 81. Lindfors K, Ciacci C, Kurppa K, Lundin KEA, Makharia GK, Mearin ML, Murray JA, Verdu EF, Kaukinen K. 2019. Coeliac disease. Nat Rev Dis Primers 5: 3
- 82. Ling HQ, Ma B, Shi X, Liu H, Dong L, Sun H, Cao Y, Gao Q, Zheng S, Li Y, Yu Y, Du H, Qi M, Li Y, Lu H, Yu H, Cui Y, Wang N, Chen C, Wu H, Zhao Y, Zhang J, Li Y, Zhou W, Zhang B, Hu W, van Eijk MJT, Tang J, Witsenboer HMA, Zhao S, Li Z, Zhang A, Wang D, Liang C. 2018. Genome sequence of the progenitor of wheat A subgenome Triticum urartu. Nature 557: 424-428.
- 83. Liu D, Yang H, Zhang Z, Chen Q, Guo W, Rossi V, Xin M, Du J, Hu Z, Liu J, Peng H, Ni Z, Sun Q, Yao Y. 2023. An elite γ-gliadin allele improves end-use quality in wheat. New Phytol 239: 5-6.
- 84. Liu J, Li D, Zhu P, Qiu S, Yao K, Zhuang Y, Chen C, Liu G, Wen M, Guo R, Yao W, Deng Y, Shen X, Li T. 2023. The landscapes of gluten regulatory network in elite wheat cultivars contrasting in gluten strength. Int J Mol Sci 24: 9447
- 85. Luo MC, Gu YQ, Puiu D, Wang H, Twardziok SO, Deal KR, Huo N, Zhu T, Wang L, Wang Y, McGuire PE, Liu S, Long H, Ramasamy RK, Rodriguez JC, Van SL, Yuan L, Wang Z, Xia Z, Xiao L, Anderson OD, Ouyang S, Liang Y, Zimin AV, Pertea G, Qi P, Bennetzen JL, Dai X, Dawson MW, Müller HG, Kugler K, Rivarola-Duarte L, Spannagl M, Mayer KFX, Lu FH, Bevan MW, Leroy P, Li P, You FM, Sun Q, Liu Z, Lyons E, Wicker T, Salzberg SL, Devos KM, Dvořák J. 2017. Genome sequence of the progenitor of the wheat D genome Aegilops tauschii. Nature 551: 498-502.
- 86. Makai S, Tamás L, Juhász A. 2014. Distinct regulatory modules identified in the promoters of wheat Glu-1 genes suggest different regulatory mechanisms. bioRxiv 19: 011635.
- 87. Makharia GK. 2015. Celiac disease screening in southern and east Asia. Dig Dis 33: 167-174.
- 88. Mangova M, Rachovska G. 2004. Technological characteristics of newly developed mutant common winter wheat lines. Plant Soil Environ 50: 84-87.
- 89. Marchioni Beery RM, Birk JW. 2015. Wheat-related disorders reviewed: making a grain of sense. Expert Rev Gastroenterol Hepatol 9: 851-864.
- 90. Marín-Sanz M, Barro F. 2022. RNAi silencing of wheat gliadins alters the network of transcription factors that regulate the synthesis of seed storage proteins toward maintaining grain protein levels. Front Plant Sci 13: 935851.
- 91. Marín-Sanz M, Iehisa JCM, Barro F. 2022. New transcriptomic insights in two RNAi wheat lines with the gliadins strongly down-regulated by two endosperm specific promoters. Crop J 10: 194-203.
- 92. Mastani S, Bahmanyar F, Shojaee-Aliabadi S, Mirmoghtadaie L, Hosseini SM. 2023. Effect of dual physical modifications on structural and functional properties of gluten and whey protein: ultrasound and microwave. Food Sci Technol Int 30: 397-406.
- 93. Matsuo H, Kohno K, Morita E. 2005. Molecular cloning, recombinant expression and IgE-binding epitope of ω-5 gliadin, a major allergen in wheat-dependent exercise-induced anaphylaxis. FEBS J 272: 4431-4438.
- 94. Matsuo H, Morita E, Tatham AS, Morimoto K, Horikawa T, Osuna H, Ikezawa Z, Kaneko S, Kohno K, Dekio S. 2004. Identification of the IgE-binding epitope in ω-5 gliadin, a major allergen in wheat-dependent exercise-induced anaphylaxis. J Biol Chem 279: 12135-12140.
- 95. McGinnis KM. 2010. RNAi for functional genomics in plants. Brief Funct Genomics 9: 111-117.
- 96. Mena M, Vicente-Carbajosa J, Schmidt RJ, Carbonero P. 1998. An endosperm-specific DOF protein from barley, highly conserved in wheat, binds to and activates transcription from the prolamin-box of a native B-hordein promoter in barley endosperm. Plant J 16: 53-62.
- 97. Merlino M, Gaudin JC, Dardevet M, Martre P, Ravel C, Boudet J. 2023. Wheat DOF transcription factors TaSAD and WPBF regulate glutenin gene expression in cooperation with SPA. PLOS ONE 18: e0287645.
- 98. Mikhaylenko GG, Czuchajowska Z, Baik BK, Kidwell KK. 2000. Environmental influences on flour composition, dough rheology, and baking quality of spring wheat. Cereal Chem 77: 507-511.
- 99. Morita E, Matsuo H, Kohno K, Yokooji T, Yano H, Endo T. 2023. A narrative mini review on current status of hypoallergenic wheat development for IgE-mediated wheat allergy, wheat-dependent exercise-induced anaphylaxis. Foods 12: 954
- 100. Nazarenko M, Izhboldin O, Liadska I, Pashchenko N. 2023. Optimal doses and concentrations of mutagens for winter wheat breeding purposes. Grain quality. Scientific Papers. Series A. Agronomy 66: 324-329.
- 101. Ng PKW, Bushuk W. 1988. Statistical relationships between high molecular weight subunits of glutenin and breadmaking quality of Canadian-grown wheats. Cereal Chem 65: 408-413.
- 102. Ni P, Zhao Y, Zhou X, Liu Z, Huang Z, Ni Z, Sun Q, Zong Y. 2023. Efficient and versatile multiplex prime editing in hexaploid wheat. Genome Biol 24: 156
- 103. Palosuo K, Alenius H, Varjonen E, Koivuluhta M, Mikkola J, Keskinen H, Kalkkinen N, Reunala T. 1999. A novel wheat gliadin as a cause of exercise-induced anaphylaxis. J Allergy Clin Immunol 103: 912-917.
- 104. Palosuo K, Varjonen E, Kekki OM, Klemola T, Kalkkinen N, Alenius H, Reunala T. 2001. Wheat omega-5 gliadin is a major allergen in children with immediate allergy to ingested wheat. J Allergy Clin Immunol 108: 634-638.
- 105. Park SC, Joung YH, Kim KM, Kim JK, Koh HJ. 2019. Gene-edited crops: present status and their future. Korean J Breed Sci 51: 175-183.
- 106. Pathirana R. 2011. Plant mutation breeding in agriculture. CABI Rev 6: 1-20.
- 107. Payne PI, Law CN, Mudd EE. 1980. Control by homologous group 1 chromosomes of the high-molecular-weight subunits of glutenin, a major protein of wheat endosperm. Theor Appl Genet 58: 113-120.
- 108. Payne PI, Nightingale MA, Krattiger AF, Holt LM. 1987. The relationship between HMW glutenin subunit composition and the bread-making quality of British-grown wheat varieties. J Sci Food Agric 40: 51-65.
- 109. Peña RJ, Trethowan R, Pfeiffer WH, van Ginkel M. 2002. Quality (end-use) improvement in wheat: compositional, genetic, and environmental factors. J Crop Prod 5: 1-37.
- 110. Pistón F, Gil-Humanes J, Barro F. 2013. Integration of promoters, inverted repeat sequences and proteomic data into a model for high silencing efficiency of coeliac disease related gliadins in bread wheat. BMC Plant Biol 13: 136
- 111. Pistón F, Gil-Humanes J, Rodríguez-Quijano M, Barro F. 2011. Down-regulating γ-gliadins in bread wheat leads to non-specific increases in other gluten proteins and has no major effect on dough gluten strength. PLOS ONE 6: e24754.
- 112. Qaim M. 2016. GM crop regulation. In: Genetically modified crops and agricultrual development. Palgrave Studies in Agricultural Economics and Food Policy. Palgrave Macmillan, New York. pp. 109-134,
- 113. Radovanovic N, Coutier S, Brown D, Humphreys DG, Lukow OM. 2002. Genetic variance for gluten strength contributed by high molecular weight glutenin proteins. Cereal Chem 79: 843-849.
- 114. Rahemi MR, Yamchi A, Navabpour S, Soltanloo H. 2015. The effect of Gamma ray on baking properties of the bread wheat Roshan. Biol Forum Int J 7: 1616-1621.
- 115. Ravel C, Fiquet S, Boudet J, Dardevet M, Vincent J, Merlino M, Michard R, Martre P. 2014. Conserved cis-regulatory modules in promoters of genes encoding wheat high-molecular-weight gluten subunits. Front Plant Sci 5: 621
- 116. Rogers WJ, Cogliatti M, Lerner SE, Ponzio NR, Robutti JL, Di Martino AM, Borrás FS, Seghezzo ML, Molfese ER. 2006. Effects of nitrogen and sulfur fertilizers on gliadin composition of several cultivars of durum wheat. Cereal Chem 83: 677-683.
- 117. Roychowdhury R, Ghatak A, Kumar M, Samantara K, Weckwerth W, Chaturvedi P. 2024. Accelerating wheat improvement through trait characterization: advances and perspectives. Physiol Plant 176: e14544.
- 118. Sánchez-León S, Gil-Humanes J, Ozuna CV, Giménez MJ, Sousa C, Voytas DF, Barro F. 2018. Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol J 16: 902-910.
- 119. Sánchez-León S, Giménez MJ, Comino I, Sousa C, Casado MÁL, Torres MI, Barro F. 2019. Stimulatory response of celiac disease peripheral blood mononuclear cells induced by RNAi wheat lines differing in grain protein composition. Nutrients 11: 2933
- 120. Sapone A, Bai JC, Ciacci C, Dolinsek J, Green PHR, Hadjivassiliou M, Kaukinen K, Rostami K, Sanders DS, Schumann M, Ullrich R, Villalta D, Volta U, Catassi C, Fasano A. 2012. Spectrum of gluten-related disorders: Consensus on new nomenclature and classification. BMC Med 10: 13
- 121. Sato S, Utsunomiya T, Imai T, Yanagida N, Asaumi T, Ogura K, Koike Y, Hayashi N, Okada Y, Shukuya A, Ebisawa M. 2015. Wheat oral immunotherapy for wheat-induced anaphylaxis. J Allergy Clin Immunol 136: 1131-1133.
- 122. Schaart JG, Salentijn EMJ, Goryunova SV, Chidzanga C, Esselink DG, Gosman N, Bentley AR, Gilissen LJWJ, Smulders MJM. 2020. Exploring the alpha-gliadin locus: The 33-mer peptide with six overlapping coeliac disease epitopes in Triticum aestivum is derived from a subgroup of Aegilops tauschii. Plant J 106: 86-94.
- 123. Scherf KA, Brockow K, Biedermann T, Koehler P, Wieser H. 2016. Wheat-dependent exercise-induced anaphylaxis. Clin Exp Allergy 46: 10-20.
- 124. Sharma N, Bhatia S, Chunduri V, Kaur S, Sharma S, Kapoor P, Kumari A, Garg M. 2020. Pathogenesis of celiac disease and other gluten related disorders in wheat and strategies for mitigating them. Front Nutr 7: 6
- 125. Shaw LM, Turner AS, Herry L, Griffiths S, Laurie DA. 2013. Mutant alleles of Photopheriod-1 in wheat (Triticum aestivum L.) that confer a late flowering phenotype in long days. PLOS ONE 8: e79459.
- 126. Shewry PR, Halford NG, Tatham AS, Popineau Y, Lafiandra D, Belton PS. 2003. The high molecular weight subunits of wheat glutenin and their role in determining wheat processing properties. Adv Food Nutr Res 45: 219-302.
- 127. Shewry PR, Halford NG, Tatham AS. 1992. High molecular weight subunits of wheat glutenin. J Cereal Sci 15: 105-120.
- 128. Shewry PR, Halford NG. 2002. Cereal seed storage proteins: structures, properties and role in grain utilization. J Exp Bot 53: 947-958.
- 129. Shewry PR, Hey SJ. 2015. The contribution of wheat to human diet and health. Food Energy Secur 4: 178-202.
- 130. Shewry PR, Lafiandra D. 2022. Wheat glutenin polymers 1. structure, assembly and properties. J Cereal Sci 106: 103486.
- 131. Shewry PR, Tatham AS, Barro F, Barcelo P, Lazzeri P. 1995. Biotechnology of breadmaking: Unreveling and manipulating the multi-protein gluten complex. Nat Biotechnol 13: 1185-1190.
- 132. Shewry PR, Tatham AS. 1997. Disulphide bonds in wheat gluten proteins. J Cereal Sci 25: 207-227.
- 133. Shewry PR. 2009. Wheat. J Exp Bot 60: 1537-1553.
- 134. Shewry PR. 2019. What is gluten - why is it special? Front Nutr 6: 101
- 135. Singh H, MacRitchie F. 2001. Application of polymer science to properties of gluten. J Cereal Sci 33: 231-243.
- 136. Singh NK, Shepherd KW. 1988. Linkage mapping of genes controlling endosperm storage proteins in wheat. Theor Appl Genet 75: 628-641.
- 137. Sissons MJ, Ames NP, Hare RA, Clarke JM. 2005. Relationship between glutenin subunit composition and gluten strength measurements in durum wheat. J Sci Food Agric 85: 2445-2452.
- 138. Sollid LM, Qiao SW, Anderson RP, Gianfrani C, Koning F. 2012. Nomenclature and listings of celiac disease relevant gluten T-cell epitopes restricted by HLA-DQ molecules. Immunogenetics 64: 455-460.
- 139. Sollid LM. 2002. Coeliac disease: dissecting a complex inflammatory disorder. Nat Rev Immunol 2: 647-655.
- 140. Tao HP, Adalsteins AE, Kasarda DD. 1992. Intermolecular disulfide bonds link specific high-molecular-weight glutenin subunits in wheat endosperm. Biochim Biophys Acta 1159: 13-21.
- 141. Taraghikhah N, Ashtari S, Asri N, Shahbazkhani B, Al-Dulaimi D, Rostami-Nejad M, Rezaei-Tavirani M, Razzaghi MR, Zali MR. 2020. An updated overview of spectrum of gluten-related disorders: clinical and diagnostic aspects. BMC Gastroenterol 20: 258
- 142. Tatham AS, Drake AF, Shewry PR. 1990. Conformational studies of synthetic peptides corresponding to the repetitive regions of the high molecular weight (HMW) glutenin subunits of wheat. J Cereal Sci 11: 189-200.
- 143. Travella S, Klimm TE, Keller B. 2006. RNA interference-based gene silencing as an efficient tool for functional genomics in hexaploid bread wheat. Plant Physiol 142: 6-20.
- 144. Tsiatsiani L, Heck AJR. 2015. Proteomics beyond trypsin. FEBS J 282: 2612-2626.
- 145. Turnbull C, Lillemo M, Hvoslef-Eide TA. 2021. Global regulation of genetically modified crops amid the gene edited crop boom - a review. Front Plant Sci 12: 630396.
- 146. kjbs-57-2-199-f JA, Stewart JA, Dromey JA, Beissbarth T, van Heel DA, Tatham A, Henderson K, Mannering SI, Gianfrani C, Jewell DP, Hill AVS, McCluskey J, Rossjohn J, Anderson RP. 2010. Comprehensive, quantitative mapping of T cell epitopes in gluten in celiac disease. Sci Transl Med 2: 41-51.
- 147. Urade R, Sato N, Sugiyama M. 2018. Gliadins from wheat grain: an overview, from primary structure to nanostructures of aggregates. Biophys Rev 10: 435-443.
- 148. Vader W, Kooy Y, van Veelen P, de Ru A, Harris D, Benckhuijsen W, Peña S, Mearin L, Drijfhout JW, Koning F. 2002. The gluten response in children with celiac disease is directed toward multiple gliadin and gluten peptides. Gastroenterology 122: 1729-1737.
- 149. Van Dijk M, Morley T, Rau ML, Saghai Y. 2021. A meta-anlaysis of projected global food demand and population at risk of hunger for the period 2010-2050. Nat Food 2: 494-501.
- 150. Verma AK, Mandal S, Tiwari A, Monachesi C, Catassi GN, Srivastava A, Gatti S, Lionetti E, Catassi C. 2021. Current status and perspectives on the application of CRISPR/Cas9 gene-editing system to develop a low-gluten, non-transgenic wheat variety. Foods 10: 2351
- 151. Walkowiak S, Gao L, Monat C, Haberer G, Kassa MT, Brinton J, Ramirez-Gonzalez RH, Kolodziej MC, Delorean E, Thambugala D, Klymiuk V, Byrns B, Gundlach H, Bandi V, Siri JN, Nilsen K, Aquino C, Himmelbach A, Copetti D, Ban T, Venturini L, Bevan M, Clavijo B, Koo DH, Ens J, Wiebe K, N'Diaye A, Fritz AK, Gutwin C, Fiebig A, Fosker C, Fu BX, Accinelli GG, Gardner KA, Fradgley N, Gutierrez-Gonzalez J, Halstead-Nussloch G, Hatakeyama M, Koh CS, Deek J, Costamagna AC, Fobert P, Heavens D, Kanamori H, Kawaura K, Kobayashi F, Krasileva K, Kuo T, McKenzie N, Murata K, Nabeka Y, Paape T, Padmarasu S, Percival-Alwyn L, Kagale S, Scholz U, Sese J, Juliana P, Singh R, Shimizu-Inatsugi R, Swarbreck D, Cockram J, Budak H, Tameshige T, Tanaka T, Tsuji H, Wright J, Wu J, Steuernagel B, Small I, Cloutier S, Keeble-Gagnère G, Muehlbauer G, Tibbets J, Nasuda S, Melonek J, Hucl PJ, Sharpe AG, Clark M, Legg E, Bharti A, Langridge P, Hall A, Uauy C, Mascher M, Krattinger SG, Handa H, Shimizu KK, Distelfeld A, Chalmers K, Keller B, Mayer KFX, Poland J, Stein N, McCartney CA, Spannagl M, Wicker T, Pozniak CJ. 2020. Multiple wheat genomes reveal global variation in modern breeding. Nature 588: 277-283.
- 152. Wang D, Li F, Cao S, Zhang K. 2020. Genomic and functional genomics analyses of gluten proteins and prospect for simultaneous improvement of end-use and health-related traits in wheat. Theor Appl Genet 133: 1521-1539.
- 153. Wang W, Pan Q, He F, Akhunova A, Chao S, Trick H, Akhunov E. 2018. Transgenerational CRISPR-Cas9 activity facilitates multiplex gene editing in allopolyploid wheat. CRISPR J 1: 65-74.
- 154. Wieser H, Koehler P, Scherf KA. 2020. Chemical composition of wheat grains. In: Wheat - An exceptional crop. Elsevier, Duxford: pp. 13-44.
- 155. Wieser H, Koehler P, Scherf KA. 2022. Chemistry of wheat gluten proteins: Quantitative composition. Cereal Chem 100: 36-55.
- 156. Wieser H, Koehler P, Scherf KA. 2023. Chemistry of wheat gluten proteins: qualtative composition. Cereal Chem 100: 23-35.
- 157. Wieser H. 2007. Chemistry of gluten proteins. Food Microbiol 24: 115-119.
- 158. Wohlers AE. 2010. Regulating genetically modified food: policy trajectories, political culture, and risk perceptions in the U.S., Canada, and EU. Politics Life Sci 29: 17-39.
- 159. Wohlers TE. In: Mercantini JM, Faucher C. 2015. The role of risk perception and political culture: A comparative study of regulating genetically modified food. (Eds). Risk and cognition. Vol. 80: Intelligent Systems Reference Library, Springer, Berlin: pp. 21-59.
- 160. Wrigley CW. 1996. Giant proteins with flour power. Nature 381: 738-739.
- 161. Xie K, Minkenberg B, Yang Y. 2015. Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. Proc. Natl Acad Sci 112: 3570-3575.
- 162. Yachha SK. 2006. Celiac disease: India on the global map. J Gastroenterol Hepatol 21: 1511-1513.
- 163. Yahata E, Maruyama-Funatsuki W, Nishio Z, Yamamoto Y, Hanaoka A, Sugiyama H, Tanida M, Saruyama H. 2006. Relationship between the dough quality and content of specific glutenin proteins in wheat mill streams, and its application to making flour suitable for instant Chinese noodles. Biosci Biotechnol Biochem 70: 788-797.
- 164. Yang T, Wang Y, Jiang J, Wang P, Zhong Y, Zhou Q, Wang X, Cai J, Huang M, Jiang D, Dai T, Cao W. 2023. Influence of high-molecular-weight glutenin subunit on components and multiscale structure of gluten and dough quality in soft wheat. J Agric Food Chem 71: 4943-4956.
- 165. Yao Y, Jia Y, Lu X, Li H. 2022. Release and conformational changes in allergenic proteins from wheat gluten induced by high hydrostatic pressure. Food Chem 368: 130805.
- 166. Yu Z, Yunusbaev U, Fritz A, Tilley M, Akhunova A, Trick H, Akhunov E. 2023. CRISPR-based editing of the ω- and γ-gliadin gene clusters reduces wheat immunoreactivity without affecting grain protein quality. Plant Biotechnol J 22: 892-903.
- 167. Zhang H, Claver IP, Zhu KX, Zhou H. 2011. The effect of ultrasound on the functional properties of wheat gluten. Molecules 16: 4231-4240.
- 168. Zhang K, Wen Q, Li T, Wang Y, Zhang Y, Luo D. 2022. Comparative study of the effects of ultrasonic power on the structure and functional properties of gliadin in wheat and green wheat. J Food Sci 87: 1020-1034.
- 169. Zhang M, Jia R, Ma M, Yang T, Sun Q, Li M. 2022. Versatile wheat gluten: functional properties and application in the food-related industry Crit. Rev. Food Sci Nutr 63: 10444-10460.
- 170. Zhao G, Zou C, Li K, Wang K, Li T, Gao L, Zhang X, Wang H, Yang Z, Liu X, Jiang W, Mao L, Kong X, Jiao Y, Jia J. 2017. The Aegilops tauschii genome reveals multiple impacts of transposons. Nat Plants 3: 946-955.
- 171. Zhou S, Zhang C, Zhao H, Lyu M, Dong F, Liu Y, Yang F, Wang H, Chai J. 2021. RNA interference targeting ω-secalin genes differentially affects the processing quality in a wheat T1BL⋅1RS translocation line. Crop J 9: 456-464.