Abstract
Gayabyeo, a Tongil-type rice variety, has been known to be resistant to the brown planthopper (BPH) in Korea. For genetic analysis of BPH resistance of Gayabyeo, we developed an F2 and F3 population derived from a cross between Gayabyeo and Taebaegbyeo which is a Tongil-type BPH susceptible rice variety. Based on the previously detected 284,501 putative SNPs between Gayabyeo and Taebaegbyeo, 99 cleaved amplified polymorphic sequences (CAPS) markers were developed, and they have been used for genotyping 180 F2 plants. By comparison of resequencing data of Gayabyeo and the sequences of already reported BPH resistance genes (Bph3, BPH9, Bph14, BPH18, BPH26), it was revealed that Gayabyeo has Bph3 and BPH26 resistance genes. Two InDel markers, Bph3IND and BPH26IND, were developed, which can be used as selection markers in breeding program aiming at introducing BPH resistance genes of Gayabyeo into Korean high quality japonica rice varieties. In addition, BPH bioassay was performed with 180 F3 lines for BPH resistance QTL analysis. Two major QTLs were found on chromosome 4 and 12. The regions of these two QTLs included Bph3 and BPH26, which also supported that Gayabyeo has Bph3 and BPH26 resistance genes. These results would be useful in accelerating development of various BPH-resistant high quality japonica rice varieties in Korea.
-
Keywords: Rice; brown planthopper (BPH); resistance; selection marker
서언
벼멸구(rice brown planthopper,
Nilaparvata lugens Stål)는 주둥이의 구침(stylets)을 이용해 주로 벼(
Oryza sativa L.)의 체관부 양액을 빨아먹는 노린재목(Hemiptera) 멸구과(Delphacidae)에 속하는 벼의 주요 해충이다(
Seo et al. 2009,
Sogawa 1982). 벼멸구는 우리나라에서 월동을 하지 못하고 매년 장시형 성충이 주로 6월 중순과 7월 하순 사이에 중국 남부로부터 기류를 타고 약 1,000 km 이상의 장거리 비행을 통해 날아와 벼에 피해를 주고 있다(
Otuka 2009,
Uhm et al. 1988). 벼의 아랫부분에 밀집 서식하면서 직접 섭식을 통해 심할 경우 벼 식물체를 고사시키는 “집중고사현상(hopperburn)”을 유발하기도 하며 rice ragged stunt virus (RRSV)와 rice grassy stunt virus (RGSV) 등의 바이러스 병을 매개하여 큰 피해가 발생한다(
Hibino 1996,
Wilson & Claridge 1991). 최근 20년동안 우리나라의 벼멸구 발생면적(ha)은 1998년(164,577), 2013년(104,599), 2005년(90,500), 그리고 2006년(62,279) 순으로 가장 넓었으며 2005년에는 집중고사 피해가 발생한 바 있다(
http://ncpms.rda.go.kr). 벼멸구는 우리나라뿐만 아니라 아시아 지역 대부분의 나라들에서 심각한 피해를 끼치고 있다. 중국과 베트남에서는 1990년대 이후 벼멸구 피해가 줄다가 2006년과 2007년에 다시 큰 피해가 발생한 바 있다(
Cantindig et al. 2009). 중국에서는 2005년에서 2007년에 걸쳐 약 25백만 ha의 지역에서 발생했으며, 베트남에서는 2002년에서 2009년 사이 매년 약 400,000 ton의 수확량 손실이 있었고, 태국에서는 2009년과 2011년 사이에 3백만 ha 이상에서 발생하여 1,100,000 ton의 수확량 손실이 발생했다(
Fujita et al. 2013).
벼멸구의 종합적 방제에 있어서 저항성 품종의 재배는 중요한 한 축이다. 살충제의 지나친 사용은 벼멸구 천적까지 사멸시켜서 오히려 벼멸구의 2차 대발생을 초래하기도 한다. 벼멸구에 대한 저항성 품종 육성 및 유전연구는 초기에 국제벼연구소(IRRI)를 중심으로 수행되었는데 1967년 벼멸구저항성 인디카 재래종인 Mudgo가 발견되고 1973년 저항성 품종인 IR26이 육성 보급된 이후 다수의 저항성 품종들이 개발되었다(
Kim et al. 1991). 우리나라에서는 1975년 이후 밀양30호를 시작으로 다수의 통일형 저항성 품종들이 육성되었으나(
Kim et al. 1991), 자포니카형 저항성 품종은 화청벼, 친농벼, 친들벼, 안미벼 등으로 매우 부족한 실정이다.
현재까지 벼멸구 저항성 인디카 재래종 품종인 Mudgo, Swarnalata, Babawee, Rathu Heenati 등과 야생벼(
Oryza officinalis, Oryza nivara, Oryza minuta, Oryza australiensis 등)으로부터 약 29개의 저항성 유전자좌들이 벼 3, 4, 6, 7, 11, 12번 염색체들에서 보고되었다(
Hu et al. 2016). 이들 중
Bph14, Bph3, BPH26, BPH18, BPH9 유전자들이 유전지도기반 유전자분리(map-based gene cloning) 연구를 통해 분리되었다.
Bph14는 3번 염색체 장완에 위치하는 야생벼
Oryza officinalis 유래 유전자로 NBS-LRR 단백질을 코딩하고 있다(
Du et al. 2009).
Bph3는 인디카 재래종 Rathu Heenati로부터 유래한 광범위 내구저항성(broad-spectrum and durable resistance)을 보이는 유전자로 4번 염색체 단완에 위치하는 세 개의 lectin receptor kinase 유전자들(OsLecRK1, OsLecRK2, OsLecRK3)로 이루어져 있다(
Liu et al. 2014).
BPH26 유전자는 인디카 품종인 ADR52에서 분리되었으며 12번 염색체 장완에 위치하고 NBS-LRR 단백질을 코딩하고 있는데
BPH2와 동일한 유전자로 밝혀졌다(
Tamura et al. 2014).
BPH18 유전자는 야생벼
Oryza australiensis의 introgression line인 인디카 계통 IR65482-7-216-1-2에서 분리되었으며 12번 염색체 장완에 위치하고 NBS-LRR 단백질을 코딩하고 있다(
Ji et al. 2016).
BPH9는 인디카 저항성 재래종인 Pokkali로부터 분리되었으며 12번 염색체 장완에 위치하고 NBS-LRR 단백질을 코딩하고 있다(
Zhao et al. 2016). 또한 12번 염색체 장완에 위치한 것으로 보고된 8개의 저항성 유전자좌들이
BPH9 유전자의 allele들인 것으로 밝혀졌으며 4종의 저항성 유전자 allele type(
BPH1/9-1, BPH1/9-2, BPH1/9-7, BPH1/9-9)으로 분류되었는데,
BPH1/9-1 allele 그룹에는
BPH1, BPH10, BPH18, BPH21 유전자들이 속하고,
BPH1/9-2 allele 그룹에는
BPH2와
BPH26 유전자들이 속하고,
BPH1/9-7 allele 그룹에는
BPH7 유전자가 속하고,
BPH1/9-9 allele 그룹에는
BPH9 유전자가 속하였다(
Zhao et al. 2016).
기존에 육성된 벼멸구 저항성 품종들 가운데 한 개의 저항성 유전자가 도입된 품종들은 보급 후 일정기간이 지난 후 저항성이 무너지는 경향을 보였다(
Hu et al. 2016). 그러나, 두 개 이상의 저항성 유전자를 집적한 계통들은 향상된 광범위 저항성을 보였다는 연구결과들이 있다(
Hu et al. 2013,
Myint et al. 2012,
Qiu et al. 2012). 그러므로, 기존에 국내 자포니카 벼 품종에 도입되지 않았던 저항성 유전자들을 도입하여 저항성 자원을 다양화하고, 몇 개의 저항성 유전자들을 집적하여 벼멸구에 대한 내구저항성을 가진 벼 품종을 육성하는 것이 필요하다.
가야벼의 벼멸구 저항성 유전자를 탐색하기 위한 선행연구로서 가야벼와 벼멸구 감수성 통일형 품종인 태백벼를 resequencing하여 두 품종간의 잠재적 SNP 약 28만개를 탐색한 바 있다(
Ji et al. 2016). 이 연구에서는 발굴된 SNP들 가운데 제한효소인식부위에 위치한 SNP들을 대상으로 새로운 CAPS(Cleaved Amplified Polymorphic Sequences) 마커 99종을 개발하였으며, 태백벼/가야벼 F2/F3 집단을 이용하여 이들 마커들의 유전지도를 작성하고, 벼멸구 저항성 QTL 분석을 수행하였다. 그리고, 가야벼의 resequencing 데이터와 기존에 밝혀진 벼멸구 저항성 유전자들의 염기서열을 비교하여, 가야벼가
Bph3와
BPH26 유전자들을 가지고 있음을 발견하였다. 또한, 가야벼를 이용한 벼멸구저항성 자포니카형 고품질벼 육종프로그램에 활용될 수 있는 가야벼의
Bph3와
BPH26 유전자 선발마커로 삽입/결실(InDel) 마커인
Bph3IND와
BPH26IND를 개발하였다.
재료 및 방법
가야벼 벼멸구 저항성 유전분석 집단의 육성
2014년 하계에 벼멸구 저항성 품종인 가야벼를 부본으로 하고 감수성 품종인 태백벼를 모본으로하여 인공교배를 수행하였고, 2014년~2015년 동계에 F1 식물체들을 온실에서 1/5000 와그너포트를 이용해서 재배하였다. F2 집단은 2015년 하계에 국립농업과학원 농업생명자원부 포장에서 30 cm × 15 cm 재식밀도로 1주 1본식으로 심어 재배하였다. 생육성기의 잎을 채취하여 Inclone plant genomic DNA prep kit를 이용하여 DNA를 추출하였다.
모본간 다형성 마커 개발과 유전지도 작성
선행연구에서 가야벼와 태백벼 유전체 서열 데이터를 이용하여 두 품종간 잠재적 SNP 약 28만개를 탐색한 바 있다(
Ji et al. 2016). CAPS 마커를 디자인하기 위해서 자체 작성한 Python 프로그램을 이용하여 이들 SNP 가운데, 비교적 흔히 사용되는 제한효소 17종(BamHI, BglII, EcoRI, EcoRV, HindIII, KpnI, Mlu I, NruI, PshBI, PvuII, SacI, SacII, SalI, ScaI, StuI, XbaI, XhoI)의 인식부위에 위치한 SNP들을 추출하였다. 그리고, 이들 SNP의 인접서열 왼쪽 500bp와 오른쪽 500bp를 추출하여 SNP를 포함한 1,001 bp의 서열에서 PCR 프라이머를 디자인하였다. 프라이머 디자인은 BatchPrimer3 1.0(
http://probes.pw.usda.gov/batchprimer3/) 프로그램을 이용해서 수행하였다. 마커와 마커 사이에 약 2 Mbp 간격을 두고 디자인하였으며, 디자인된 마커로 가야벼와 태백벼의 DNA로 모본간 다형성 검정을 수행하여 실제로 다형성을 보이는 마커들을 선발하였다. 선발된 CAPS 마커들로 F2 집단의 개체별 유전자형 분석을 실시하였다. CAPS 마커 분석의 PCR 조건은 94°C 3분 후 94°C 40초, 60°C 40초, 72°C 1분40초의 35 사이클, 그리고 72°C 7분이었다. PCR 산물을 제한효소로 37°C에서 8시간 이상 절단한 후 1.2% 아가로스젤에서 전기영동하였다. 이렇게 생산된 F2 집단의 개체별 유전자형 데이터를 가지고 MapDisto 1.7 프로그램(
Lorieux 2012)을 이용하여 마커들의 유전지도를 작성하였다.
기존에 밝혀진 벼멸구저항성 유전자인
Bph14, Bph3, BPH26, BPH18, BPH9들의 염기서열을 reference 염기서열로 하고, 선행연구(
Ji et al. 2016)에서 Illumina Hiseq2000 장비를 이용하여 얻어진 가야벼의 유전체 염기서열들을 reference 염기서열에 read mapping하였다. read mapping은 CLC Assembly Cell 프로그램(ver. 3.2.2,
http://www.clcbio.com)의 clc_mapper 명령어를 사용하여 수행하였는데, 이 때 옵션 값을 similarity 98%, matched length fraction 1.0, repeat ignore로 설정하여 각 read의 전체 영역이 match되면서 염기서열 유사도가 98% 이상인 서열만이 reference 염기서열에 mapping 되도록 하고 반복서열들은 제외되도록 하였다. read mapping 결과를 clc_mapping_viewer 명령어를 시용하여 검토함으로써 가야벼의 유전체 염기서열이 비교대상인 각각의 벼멸구저항성 유전자 염기서열과 일치하는지를 판정하였다. 이 결과, 가야벼는
Bph3 유전자와
BPH26 유전자를 가지고 있는 것으로 나타났다. 이를 더 검증하기 위해 가야벼의
Bph3 유전자와
BPH26 영역을 PCR로 증폭하여 Sanger sequencing 방법으로 sequencing 하였다. 이 때 사용한 PCR 프라이머 목록은
Table 1과 같다.
Bph3 유전자는 세 개의 lectin receptor kinase 유전자들인 OsLecRK1, OsLecRK2, OsLecRK3로 이루어져 있는데, 이들 각각의 유전자 코딩 시퀀스 영역이 포함되도록 PCR 증폭하여 PCR 산물을 Sanger sequencing 방법으로 sequencing 하였다. 시퀀싱 프라이머는 약 500 bp 간격으로 디자인하여 제작하였다.
BPH26 유전자는 크기가 10.6 kbp로 세 개의 엑손(exon)과 두 개의 인트론(intron)으로 이루어져 있는데
Tamura et al. (2014)의 방법으로 엑손 영역만을 PCR 증폭하여 PCR 산물을 Sanger sequencing 방법으로 sequencing 하였다.
Table 1The list of primers for amplification of Bph3 and BPH26 genes of Gayabyeo
Table 1
|
Gene |
forward primer |
reverse primer |
gene size (bp) |
|
OsLecRK1 |
CCCACTCAATTATAGCTACACA |
TCGACGACAGCAAATCCT |
2442 |
|
OsLecRK2 |
ACAAATGCCGTTCTCCAAG |
AGGATCATGTGGTTGTAGAGG |
2436 |
|
OsLecRK3 |
ACGCGGTAGGAAAGACCA |
AAGCATCAGTGCCCGAACA |
2436 |
|
BPH26 exon1 |
TAGCATCAGTCCCTTGCTTGTTTGC |
ATTGATTTAATTAGCAGACAAGTTG |
1089 |
|
BPH26 exon2 |
AACTCTCGTCTCGTCTTAAAATATA |
AGTAGTAATGTGCGTAGCAATGGAG |
624 |
|
BPH26 exon3 |
CTAGTGCCAGTTACTCCGATAAATAT |
TATGCACTAGCATCACTACA |
1944 |
가야벼 저항성 유전자 선발마커 개발
가야벼의
Bph3 유전자와
BPH26 유전자 선발마커를 다음과 같은 방법으로 개발하였다. 가야벼의 유전체 염기서열 데이터를 가지고 CLC Assembly Cell 프로그램의 clc_assembler 명령어를 사용하여 de novo assembly를 수행하였다. 그 결과 생산된 contig 염기서열들을 CLC Genomics Workbench 프로그램(ver. 6.01,
http://www.clcbio.com)을 이용하여 BLAST 데이터베이스로 만들고,
Bph3와
BPH26 유전자 염기서열로 BLAST를 수행하여 이들 유전자를 포함하는 가야벼 contig들을 추출하였다. 이렇게 추출한 가야벼 contig 염기서열과 자포니카 벼 표준유전체 염기서열인 니폰바레 염기서열(Nipponbare IRGSP-1.0 sequence,
http://rapdb.dna.affrc.go.jp/download/irgsp1.html)을 비교하여 삽입/결실(InDel)부위를 찾아내었다. 이들 InDel 부위 양측 인접염기서열을 이용하여 프라이머를 디자인해서 InDel 마커를 개발하였다.
태백벼/가야벼 F2 집단에서 채종한 F3 종자를 이용해서 F3 집단 180 계통에 대한 벼멸구저항성유묘검정을 수행하였다. F3 한 계통당 15립을 파종하여 유묘가 3엽기에 이르렀을 때 2~3령의 벼멸구 유충을 식물체 개체당 10~15마리 되도록 접종하였다. 감수성 대비 품종인 태백벼가 거의 완전히 고사하였을 때 식물체 개체의 저항성지수를 Huang 등에 의해 보고된 기준에 따라 0~9 등급으로 판정하였다(
Huang et al. 2001). 개발된 CAPS 마커들 및
Bph3와
BPH26 유전자 선발마커들의 유전지도와 F3 계통별 저항성지수의 평균값을 QTL 분석에 사용하였다. QTL 분석은 Windows QTL Cartographer version 2.5 program(
Basten et al. 1996)을 이용하여 composite interval mapping 방법으로 수행하였다. LOD threshold는 확률 수준 0.05에서 1,000번의 permutation을 함으로써 산정되었다.
결과 및 고찰
가야벼와 태백벼간 다형성 SNP 마커 개발
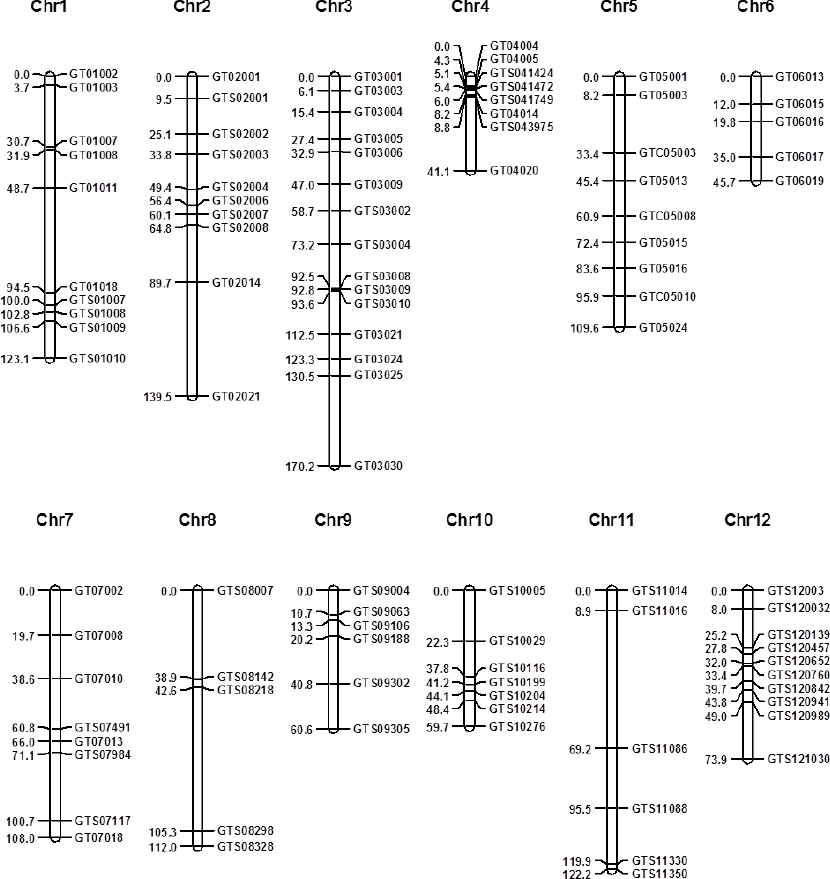
선행연구에서 탐지된 가야벼와 태백벼간 SNP들 가운데 이 연구에서 사용한 제한효소들의 인식부위에 있는 SNP들을 이용하여 CAPS 마커들을 디자인하고 가야벼와 태백벼간 다형성 검정을 하였다. 약 2 Mbp 간격으로 마커들을 디자인 하였으며, 가야벼와 태백벼간 SNP가 드물어서 마커 디자인이 불가능한 영역들도 있었다. 디자인된 마커들로 가야벼와 태백벼 DNA를 이용하여 모본 다형성 검정을 수행하였으며, 모본 품종간 다형성이 확인된 마커들로 태백벼/가야벼 후대 F2 집단 180 개체의 유전자형을 분석하였다. 99 개의 CAPS 마커들이 개발되어, 이들을 이용한 태백벼/가야벼 후대 F2 집단 유전자형 데이터들이 생산되었으며, 이 데이터에 기반하여 개발된 마커들의 유전지도를 작성하였다(
Fig. 1). 작성된 유전지도의 총 거리는 1165.5 cM이었고, 마커간 평균 거리는 13.4 cM이었다. 개발된 마커들의 프라이머 염기서열, 제한효소, 물리적 위치를
Table 2에 나타내었다.
Fig. 1Genetic map of developed CAPS markers.
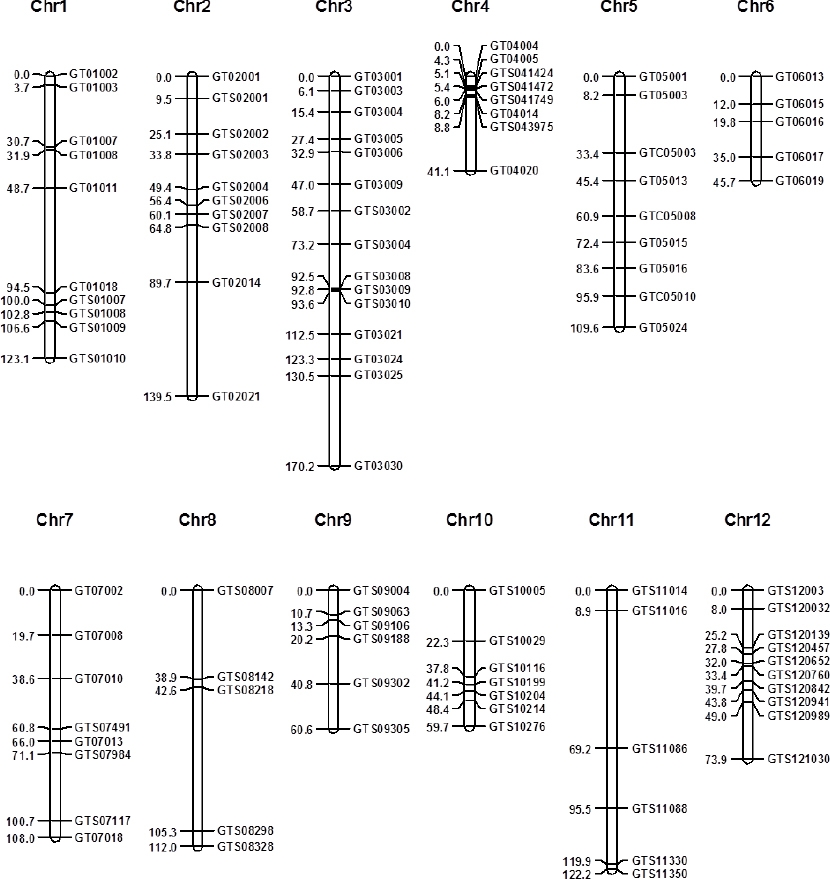
Table 2List of developed CAPS markers
Table 2
|
no. |
marker name |
forward primer |
reverse primer |
restriction enzyme |
chromosome |
position (bp) |
|
1 |
GT01002 |
GCTGATGATCGTTTGCCACA |
TGCGATGTATTTTCGCGTTTT |
EcoRV |
1 |
1,065,395 |
|
2 |
GT01003 |
AAATTGGCTGGGGCACTTCT |
TGCAGGATCAGGAAGAGCTG |
EcoRI |
1 |
1,600,379 |
|
3 |
GT01007 |
CAGGGCCGCAGTGTTAATTC |
AGCGAGACGTTCGTGATGGT |
EcoRI |
1 |
5,522,341 |
|
4 |
GT01008 |
AATAAGCGCTTGACGCGAAA |
CGCTCAACCACCTCGTCATT |
EcoRV |
1 |
6,037,347 |
|
5 |
GT01011 |
TCCTGAGCATCCTTGCATGA |
AAAACGAGAGGGCGCATGT |
EcoRV |
1 |
9,187,583 |
|
6 |
GT01018 |
ACTTGGAGGCGCATAACGAA |
GCATAATGACATGTCGCATTGAA |
EcoRI |
1 |
23,956,461 |
|
7 |
GTS01007 |
AAGGGTTGTATGAACCGTCAAC |
TCCATGGGACACTAAAGAGAGG |
SalI |
1 |
25,601,011 |
|
8 |
GTS01008 |
CGTCGAAGAAAGTGACTCAAGA |
CCTCTTTTGTTGGTATCATTGG |
NruI |
1 |
26,694,741 |
|
9 |
GTS01009 |
ACTCCCAAAACCAGCGTAAATA |
CGTCAGGAATGAATTCGAAACT |
SacI |
1 |
27,335,280 |
|
10 |
GTS01010 |
AGAACCCGTGTAGAGCACATTA |
CAATTGATCTGCTTGCTCTCTG |
PshBI |
1 |
32,523,034 |
|
11 |
GT02001 |
GCATATGTTTGACCGTTCGTCTT |
CGGCGCTACACCATACCAAT |
EcoRI |
2 |
198,216 |
|
12 |
GTS02001 |
AACTAGCACGCTTCGTTTTAGC |
CGGATTGGTGGAATTGAAAA |
StuI |
2 |
1,330,779 |
|
13 |
GTS02002 |
CCATTTGGCAATGTTCTCATAC |
CACCCTTCTATGCTCTCATCCT |
BamHI |
2 |
4,001,036 |
|
14 |
GTS02003 |
TGTGTGGTAGAAAATGGACCTG |
CCATGATTGCCAGTGTTACATC |
EcoRI |
2 |
5,066,630 |
|
15 |
GTS02004 |
TACCGATCAGCTGAAAACACAC |
TCTTGTTCCCTTCCCTTTAACA |
BamHI |
2 |
6,854,129 |
|
16 |
GTS02006 |
GTGACCTCCGATCAAATCAACT |
CTACTGTGGGCAGTGAAGAGGT |
BamHI |
2 |
8,587,683 |
|
17 |
GTS02007 |
ATTTCCCTCCTGTTCCTCATTT |
GGTTGGGTTTTGGTATAGCTGA |
HindIII |
2 |
9,657,613 |
|
18 |
GTS02008 |
AGCTGGAAAATGTTGTATGCAG |
GGGATGAATCAGTAGTTCAGCA |
HindIII |
2 |
10,645,645 |
|
19 |
GT02014 |
GTGTTGGCTAGGGCCTTTGA |
TTCTCTGGTTGGCCTTCAGC |
EcoRI |
2 |
21,056,727 |
|
20 |
GT02021 |
CGCCTCTTGAAGACCCAGAG |
CTGACCGGCATGGGAGTTTA |
SacI |
2 |
33,887,583 |
|
21 |
GT03001 |
GCTGTTGCCGCAATATCACA |
AGGCATTGGCATTGTTAGGC |
EcoRI |
3 |
855,210 |
|
22 |
GT03003 |
TGGGGATTGCCAGATGTGAT |
GCATGGCATTTGAAAGAGTGA |
PvuII |
3 |
2,538,138 |
|
23 |
GT03004 |
CAGTGTCTGGGCACAGCAAG |
GCGTTCCACACTTCCACTCC |
HindIII |
3 |
3,811,855 |
|
24 |
GT03005 |
GGAAATACCTGCCCCGTTTT |
CAGACGATCGGGAATTTCTACG |
EcoRV |
3 |
4,753,112 |
|
25 |
GT03006 |
CTCCTCTCGCACGCAGAAGT |
CCAGCTTCTCCACGGAACAA |
EcoRV |
3 |
5,592,985 |
|
26 |
GT03009 |
TCTACCTGCCCAGGAAGCAA |
TTTCAGGCAAAGCAAAGCAG |
EcoRV |
3 |
9,205,476 |
|
27 |
GTS03002 |
AGCATCACGATGATTCTTTTGC |
TTATTGTGCCCTAGACGAATGA |
SacI |
3 |
11,547,474 |
|
28 |
GTS03004 |
TAGGGTTGGCTCTCAATCTCTC |
ACACCTACAAGGTTGTGACTGC |
EcoRI |
3 |
13,593,894 |
|
29 |
GTS03008 |
ATCGCACGGAAGGATAACTG |
GTTGCTCTAGAACCTCGCAATC |
SacII |
3 |
19,416,808 |
|
30 |
GTS03009 |
ATTATCTCACGACCTGGGACTG |
AGCTAGCTAGGGTTTGTGTTGC |
EcoRI |
3 |
20,345,341 |
|
31 |
GTS03010 |
TGAGGAAGAGGAGGAGATTGAG |
CTGCTTCATTGTGTCTGAGGAC |
SacI |
3 |
21,733,967 |
|
32 |
GT03021 |
GAATCTGATTGCCCCCTAGC |
GGACCCAAATTGAAAGGCAAT |
EcoRV |
3 |
25,459,877 |
|
33 |
GT03024 |
TTGGAAGCTTGAAACGATGG |
TCGCAAGTTCGAAACCCACT |
EcoRI |
3 |
28,056,489 |
|
34 |
GT03025 |
TGCGAATACAATTCGGACTGC |
CGTGCAGTACCCTCCAAACC |
EcoRV |
3 |
29,184,626 |
|
35 |
GT03030 |
AATTAAACTCCGCGCCTGGT |
AAACCATGCATCCGATAGGG |
HindIII |
3 |
36,245,706 |
|
36 |
GT04004 |
TGGCGACAAACAGAGCACAC |
CCTTCTTGCGCAGGACCTC |
EcoRI |
4 |
4,875,618 |
|
37 |
GT04005 |
CCAGCCAAACTCAGTTGTCCA |
CAGACCATACCTCCTTCAGAGCA |
EcoRI |
4 |
5,924,429 |
|
38 |
GTS041424 |
ACTGGGAAGAAGGCAAGACA |
GTGAGCATCCCACTCTCCAT |
EcoRI |
4 |
6,623,655 |
|
39 |
GTS041472 |
CCTGAAAGGGCTCAAAAACA |
TCTTGCAGGTGACAAATCCA |
PshBI |
4 |
6,914,922 |
|
40 |
GTS041749 |
TGAACTATTGCGGTTGACCA |
TGCGGATTGTGTAAGACAGG |
EcoRV |
4 |
8,481,173 |
|
41 |
GT04014 |
TGGATCGCTTGATGTGGTTG |
ATGCCTCAAATACCGCAGCA |
EcoRI |
4 |
14,685,347 |
|
42 |
GTS043975 |
TTCGAGGGTTGGTGACTGAT |
AGAGGGTCCATGGTTTTGTG |
EcoRV |
4 |
16,607,468 |
|
43 |
GT04020 |
TCCTTGCAAACGCAAAGTCA |
GCAACAAAACATTGCGAAGG |
EcoRI |
4 |
21,083,274 |
|
44 |
GT05001 |
TTGCAGCTGAAGAAAATGTG |
TTTGGCACATGAAAATGATG |
EcoRI |
5 |
602,152 |
|
45 |
GT05003 |
TGTTGCAGAGTAGCAAACCA |
TATAAGCTGCAGCCTGGAAT |
EcoRV |
5 |
2,107,453 |
|
46 |
GTC05003 |
CCGAAGAGCGACTCCTAGTG |
AGAGATGCACAGAGGGAGGA |
PshBI |
5 |
6,043,943 |
|
47 |
GT05013 |
GAGCATAAGGGGTTTCCCTA |
TAATGCTGGTGTGGACTCAA |
EcoRI |
5 |
15,946,504 |
|
48 |
GTC05008 |
TAATTGCCTCCCAAACAACC |
TGTCATGCAAAGCAAAAAGG |
PshBI |
5 |
18,530,587 |
|
49 |
GT05015 |
CGACGACTATATGGCGGATA |
AACTCCAATCGGTCATACCA |
EcoRI |
5 |
20,548,891 |
|
50 |
GT05016 |
CCGTTTTGTCAGAAACCTGT |
TAAATCGCGAGACGAATCTT |
EcoRV |
5 |
22,331,098 |
|
51 |
GTC05010 |
AGGTGGTGGTGGTTCCATT |
TCACTGCCGATTTCTCACTG |
PvuII |
5 |
25,145,098 |
|
52 |
GT05024 |
TGATCCCCAATGAAAGCTAC |
TCTTGTACTCCCTTCGTTTCA |
EcoRI |
5 |
29,110,069 |
|
53 |
GT06013 |
TCCAACAGCAACAATCTTCA |
TCTCAAGCTCAATTCCGAGT |
EcoRI |
6 |
24,403,695 |
|
54 |
GT06015 |
AGAGCAGAGCTCGTGTTAGTG |
TGGTGATTTTTGGATGGATT |
EcoRI |
6 |
26,356,825 |
|
55 |
GT06016 |
GCGAAGATGCTAGTGGAAAA |
GTTGCAAAGGATCTGGAAAA |
EcoRV |
6 |
27,324,979 |
|
56 |
GT06017 |
GATTCGACGTCTGAAAAGGA |
ATATGCATGGTTCACTGCAA |
EcoRI |
6 |
29,168,500 |
|
57 |
GT06019 |
TGCTCAGCAGAGTCTTGCTAT |
GCTTGAGACTGGTTCGTGTT |
SacII |
6 |
31,015,468 |
|
58 |
GT07002 |
CAAAAGTAACCAATGGCAAGA |
GCACATTTGAACTGCATCTG |
EcoRV |
7 |
5,619,862 |
|
59 |
GT07008 |
TGGAATCAAAGTGAAGGTCAG |
TGGCAATCTAAAGAGGAGGA |
EcoRI |
7 |
17,797,502 |
|
60 |
GT07010 |
TATGGGGACTCCAATAAGCA |
TACGGTCTCGTCGGTATCAT |
EcoRV |
7 |
19,489,327 |
|
61 |
GTS07491 |
AAGGCCAAATGGAGGTTAGC |
CTGGGAGTCCAGTCATCACA |
BglII |
7 |
21,220,123 |
|
62 |
GT07013 |
ATTCAGCCCCTTCTGGATAC |
ACCTAGGGCAGTCCTTGATT |
EcoRI |
7 |
22,517,510 |
|
63 |
GTS07984 |
GCCGCTTTCTAAAAGGATTT |
GACCGAAATTTTGGCTCAAT |
EcoRV |
7 |
23,387,924 |
|
64 |
GTS07117 |
AGCCTGCCACTCTTCCTGTA |
CGAGGACATGGCATCCTATT |
EcoRV |
7 |
27,812,959 |
|
65 |
GT07018 |
TTCAACCTTAGGCACTCGTC |
TGAGATTCTCCTTGCGTGTT |
EcoRV |
7 |
28,923,650 |
|
66 |
GTS08007 |
GATCAGTCGTCCTCGTCCTC |
GCATTGAGCACATCCATGAC |
HindIII |
8 |
1,150,395 |
|
67 |
GTS08142 |
ATGCTCCAATTGAGCTGACC |
TACTCATGCAGGAACGCATC |
EcoRI |
8 |
6,123,634 |
|
68 |
GTS08218 |
AAATGGTGTTTTTCCCGTTG |
AGGTCGTGGCATGATTTAGC |
PshBI |
8 |
8,787,812 |
|
69 |
GTS08298 |
CGATCGGCTCTTACATGACA |
AGTGGATTTGCAGGCAGCTA |
EcoRI |
8 |
27,252,203 |
|
70 |
GTS08328 |
GAGTTACAAACCGCCTTTGC |
TTGCTAGGGGTCAAGGAAAA |
BglII |
8 |
28,243,051 |
|
71 |
GTS09004 |
AGCACCCAGAGCAACAGTTT |
CCCACTAATTAGCCCATTGC |
EcoRV |
9 |
620,373 |
|
72 |
GTS09063 |
GCTTGGATGACAAGCGTTCT |
AGATCGGCTTCCGAAAGAGT |
BglII |
9 |
9,835,918 |
|
73 |
GTS09106 |
ACGCGACTGCTAAGGTTCAT |
CTCGATCGGTCCCTTTCAT |
EcoRV |
9 |
11,310,635 |
|
74 |
GTS09188 |
AGTGGTAGATCCCGTTGAGC |
AACCGGAGACCTTCAGTGTG |
EcoRV |
9 |
13,099,244 |
|
75 |
GTS09302 |
GCATGCATCCGTTGAATAAA |
GAGATGCTCGTCAGGGACAC |
HindIII |
9 |
16,712,838 |
|
76 |
GTS09305 |
ATGGCTTTCGGGCATAGTAA |
GGATGGCCTATTCAGCAAAA |
EcoRV |
9 |
21,161,450 |
|
77 |
GTS10005 |
TCTCAGCCACTCGTTGAATG |
TCTGGATGGGCTAAATGAGG |
EcoRV |
10 |
721,349 |
|
78 |
GTS10029 |
CCTCCCTTCAGTTTGTGAGC |
CATTGGATCCCCAAAGAAAA |
EcoRI |
10 |
3,044,726 |
|
79 |
GTS10116 |
CCCCCTAAGATGGTTTTTGC |
TGAGAAGTGAAGTGTGAGTGTGG |
EcoRV |
10 |
4,577,975 |
|
80 |
GTS10199 |
AGCCTGAGAAGCAGATTGGA |
TGCAATGACGTGATGGTACA |
EcoRI |
10 |
6,149,421 |
|
81 |
GTS10204 |
TTTCACACGGGACATTCAAA |
TCGACCTTGGACTTCTAGGC |
StuI |
10 |
9,961,760 |
|
82 |
GTS10214 |
TAGGATCTCGCCGAACATCT |
ATGCACCTCCCCAACATATC |
EcoRV |
10 |
11,290,285 |
|
83 |
GTS10276 |
TGGTCTTTGAAGCATTCTGG |
TTCAATATTGGCCGTAAACC |
EcoRV |
10 |
13,637,268 |
|
84 |
GTS11014 |
CTTCGCTTCATTTCGAGTTTG |
CATTTCTGCAGTCGCATTTG |
EcoRV |
11 |
353,397 |
|
85 |
GTS11016 |
CGCACGAGATGCTGTCTAAC |
GAAAAACCCTCTGCCCTCTT |
NruI |
11 |
1,389,204 |
|
86 |
GTS11086 |
TGCTGGTGTGCTATTTGAGC |
CGTGGCCATCCAAAAGTAAT |
EcoRI |
11 |
17,330,622 |
|
87 |
GTS11088 |
CGCCTAGGTGCAGAAAAGAG |
ACACCATGAGCCTCCTCACT |
BamHI |
11 |
20,838,960 |
|
88 |
GTS11330 |
TGTCATTGGTTGCCCTGTAA |
AGCATGGAGGTGGTGCTAAC |
EcoRV |
11 |
26,534,991 |
|
89 |
GTS11350 |
AGGAATATGAATCGCGTGTG |
TTTGGGAGCTATGGCCTATG |
EcoRI |
11 |
27,197,820 |
|
90 |
GTS120003 |
GCGTTTTTCCTCCAATTCAA |
ACCGACAACTTTTGCCCTTT |
PvuII |
12 |
3,007,338 |
|
91 |
GTS120032 |
AACGAAGAGGCCAATGGATA |
GTTGACCATGGGTGCTCACT |
PvuII |
12 |
3,807,192 |
|
92 |
GTS120139 |
TGCTTTCAGGAGAGCAGGAT |
CCACCAGCATCACTCTGCTA |
EcoRI |
12 |
9,263,934 |
|
93 |
GTS120457 |
GCATGAGAGACAGCGGAGTT |
GTGATTTGCCATGCCTTTTT |
EcoRV |
12 |
13,674,598 |
|
94 |
GTS120652 |
TGGGCACAACTACAAAGGTG |
CATTCCCATGTTCCACATCA |
EcoRI |
12 |
17,438,893 |
|
95 |
GTS120760 |
CCCAGCCACAAGAAATGAAT |
TTTTCCTTTTCCCTGCTGTG |
EcoRI |
12 |
19,070,762 |
|
96 |
GTS120842 |
AAGCCATCATGTTCCATTCC |
TTTTGGTACGTCATCCGTGT |
EcoRI |
12 |
21,522,146 |
|
97 |
GTS120941 |
CAGGTGGTCCTTTTCAGCAT |
TAGTGTCATGTGCCCTGAGC |
EcoRI |
12 |
22,565,939 |
|
98 |
GTS120989 |
CCTTTGGAGGGCTTGACATA |
TACCCACTGGAAACGGAAGA |
EcoRV |
12 |
23,622,982 |
|
99 |
GTS121030 |
CGTTGCTCCGTTCATCTTCT |
GCTAGCCCCTTTTCATCTCC |
EcoRV |
12 |
26,276,873 |
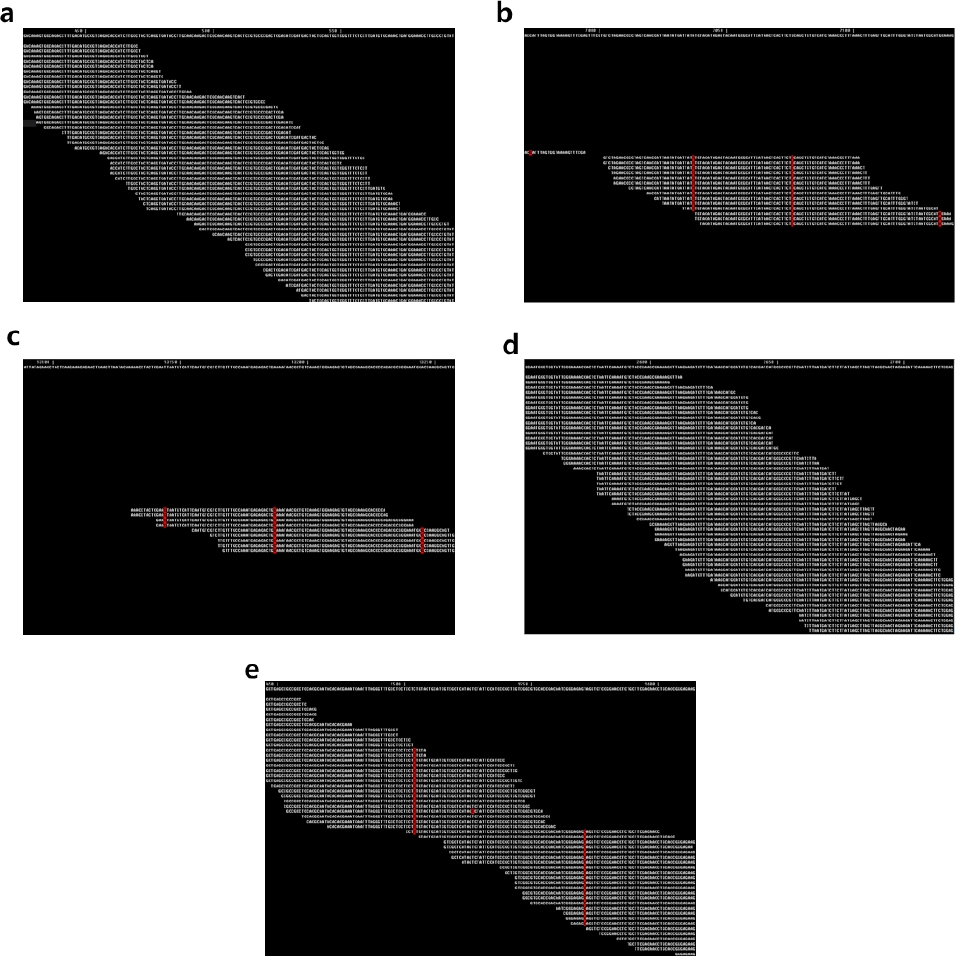
유전체 염기서열 데이터 분석에 의한 가야벼 벼멸구 저항성 유전자 규명
기존에 밝혀진 벼멸구저항성 유전자인
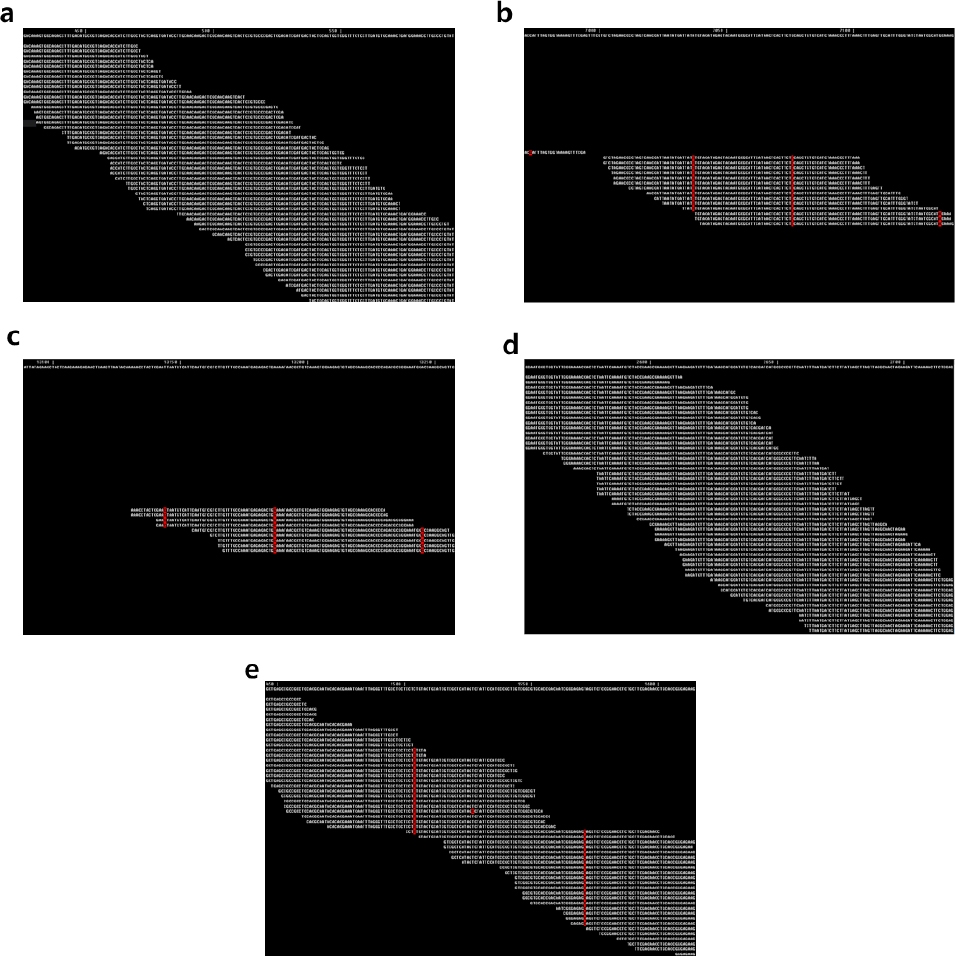
Bph14, Bph3, BPH26, BPH18, BPH9들의 염기서열을 reference 염기서열로 하여, 가야벼의 유전체 염기서열을 read mapping하여 비교한 결과, 가야벼의 유전체 염기서열이
Bph3과
BPH26 유전자 염기서열과 일치하였다(
Fig. 2).
Fig. 2에서 reference 염기서열과 일치하는 유전체 염기서열은 흰색으로 나타나고, 일치하지 않는 염기서열은 적색으로 나타나는데,
Bph3과
BPH26 유전자에서는 전 영역에 걸쳐 염기서열이 일치한 반면
Bph14, BPH18, BPH9 유전자에서는 일치하지 않는 염기서열이 다수 존재하였고, read mapping이 되지 않는 영역이 존재하였다. 따라서, 가야벼가
Bph3과
BPH26 저항성 유전자를 가지고 있다고 판단되었다.
Fig. 2
Read mapping of Gayabyeo genome sequences onto BPH resistance genes.
Reference BPH resistance gene sequences were shown at the top row of each panel, and mapped read sequences were shown below the reference sequences. White letters indicate identical nucleotides while red letters indicate different nucleotides. a: Bph3, b: BPH14, c: BPH18, d: BPH26, e: BPH9.
이 결과를 검증하기 위해 가야벼의
Bph3과
BPH26 유전자 영역을
Table 1의 프라이머로 PCR 증폭하여 Sanger sequencing 방법으로 sequencing하여
Bph3과
BPH26 유전자 염기서열과 비교하였다.
Bph3 유전자를 구성하는 세 개의 lectin receptor kinase 유전자들인 OsLecRK1, OsLecRK2, OsLecRK3 각각의 가야벼 PCR 산물의 염기서열이
Bph3 유전자 염기서열과 완전히 일치하였다. 그리고, 가야벼의
BPH26 유전자 세 개 엑손 영역 PCR 산물의 염기서열도
BPH26 유전자 영역 염기서열과 완전히 일치하였다. 이로써, 가야벼가
Bph3과
BPH26 저항성 유전자를 가지고 있음을 확인할 수 있었다.
가야벼가 벼멸구 저항성 유전자
Bph3과
BPH26을 가지고 있으므로, 이들 유전자들을 국내 자포니카형 벼 품종들에 도입하는 육종프로그램에 사용할 수 있는 선발마커를 개발하였다. 특히, 일반적인 실험실에서 아가로스 젤을 이용하여 쉽게 분석할 수 있는 삽입/결실(InDel) 마커를 개발하고자 하였다. 가야벼의 유전체 염기서열과 자포니카형 벼 품종들의 유전체 염기서열간 InDel 부위를 찾기 위해서, 먼저 가야벼의 유전체 염기서열 데이터로 de novo assembly를 수행하여 contig 들을 만들었다. de novo assembly 결과 200~95,218 bp 범위의 크기를 가진 contig들이 64,373개 생성되었으며, BLAST를 통해서
Bph3유전자와
BPH26 유전자 영역에 있는 가야벼 contig들을 추출하였다. 추출된 가야벼 contig들의 sequence와 자포니카 벼 표준유전체 염기서열인 니폰바레 염기서열과의 비교를 통해서 InDel 부위를 발굴하였다.
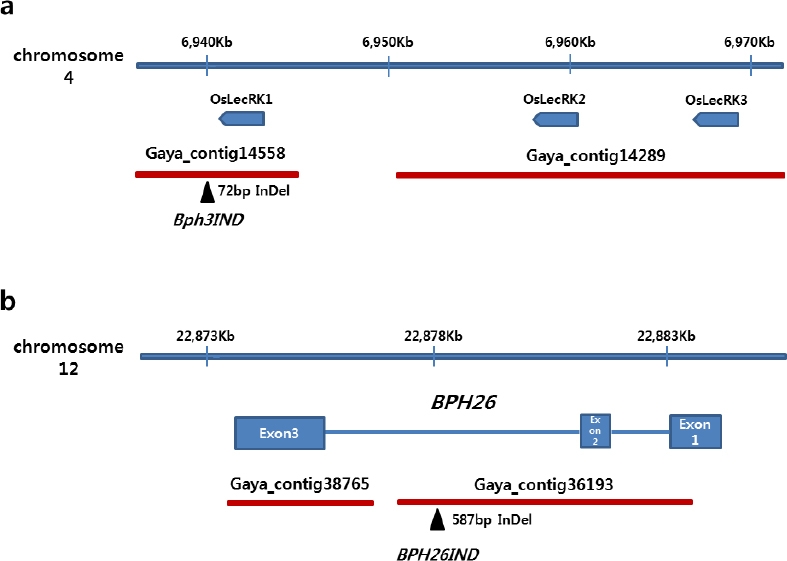
Bph3유전자를 구성하고 있는 OsLecRK1, OsLecRK2, OsLecRK3 유전자 영역에서 OsLecRK1 유전자의 3’ 말단 인접부위에서 가야벼 contig 14558과 니폰바레 유전체 염기서열간 72 bp InDel 부위를 발굴하고(
Fig. 3a) InDel 마커
Bph3IND를 개발하였다(
Table 3).
BPH26유전자는 세개의 exon과 두 개의 intron으로 구성되어 있는데 두번째 intron에서 가야벼 contig 36193과 니폰바레 유전체 염기서열간 587 bp InDel 부위를 발굴하고(
Fig. 3b) InDel 마커
Bph26IND를 개발하였다(
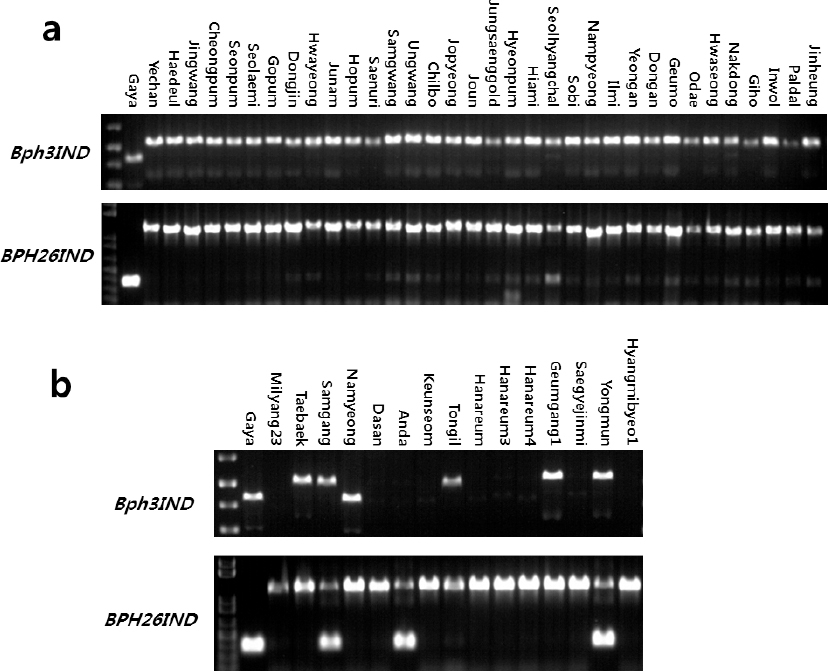
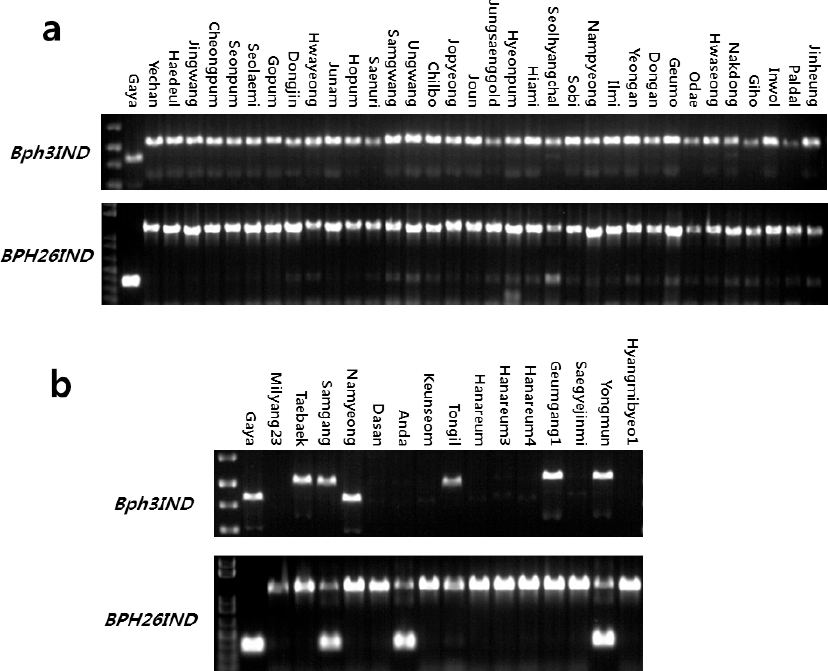
Table 3). 이들 마커들로 가야벼와 34개의 국내육성 자포니카 벼 품종들의 유전자형을 분석한 결과,
Bph3IND 마커와
Bph26IND 마커 둘 다 가야벼와 국내 자포니카 품종들간에 뚜렷한 차이를 보였다(
Fig. 4a). 그러므로 이 마커들은 가야벼의 벼멸구 저항성 유전자들을 국내 자포니카 벼 품종들에 도입하는 여교배 육종 프로그램에 선발마커로 유용하게 사용될 수 있을 것으로 판단된다. 한편,
Bph3IND 마커로 가야벼와 15개의 국내육성 통일형 벼 품종들의 유전자형을 분석한 결과,
Bph3IND 마커에서는 남영벼만이 가야벼와 동일한 밴드를 보였고, 태백벼 등 5품종에서 가야벼와 다른 밴드가 관찰되었으며, 밀양23호 등 9 품종에서는 밴드가 관찰되지 않았다(
Fig. 4b). 이로써 벼멸구 저항성 품종으로 알려진 남영벼가
Bph3 유전자를 가지고 있을 가능성이 있다고 보여진다. 또한,
Bph26IND 마커로 가야벼와 15개의 국내육성 통일형 벼 품종들의 유전자형을 분석한 결과, 밀양23호 등 12 품종에서는 가야벼와 다른 밴드가 관찰되었고, 삼강벼, 안다벼, 용문벼에서는 가야벼와 동일한 밴드가 관찰되었으나 밴드가 두 개인 hetero type들이어서 가야벼와 다른 양상을 보였다. 이로써, 국내육성 통일형 벼 품종들에서도
Bph3IND 마커와
Bph26IND 마커의 활용가능성이 있다고 판단된다.
Fig. 3Detection of InDels between Gayabyeo and Nipponbare reference sequences for development of InDel markes in Bph3 and BPH26 region. a: Bph3 region, b: BPH26 region.
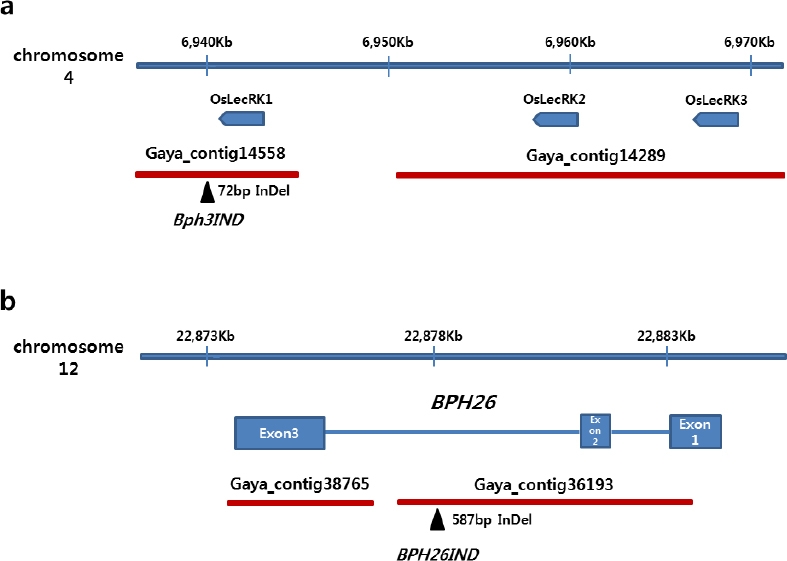
Table 3Primer sequences of Bph3IND and BPH26IND markers.
Table 3
|
marker name |
forward primer |
reverse primer |
|
Bph3IND
|
GCTTCACCTGTGTCCTGTT |
GTGTATGAGGGTTGTTCTGGAT |
|
BPH26IND
|
GGGCAGCTAAGCGCATGGTT |
GGGTGATAAATCGAACAGCAGAGG |
Fig. 4
Genotyping of Gayabyeo and other Korean rice varieties with Bph3IND and BPH26IND markers.
a: genotyping Gayabyeo and 34 Korean japonica rice varieties, b: genotyping Gayabyeo and 15 Korean Tongil-type rice varieties.
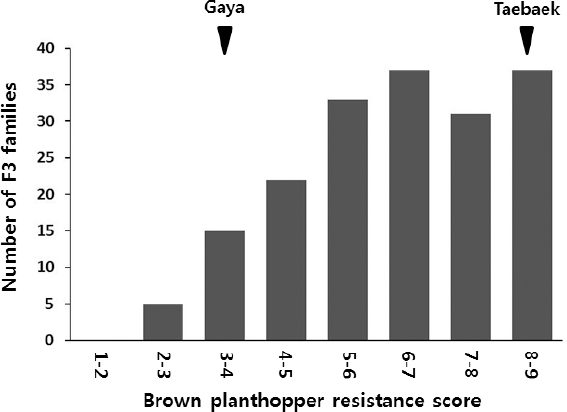
가야벼의 벼멸구저항성 QTL mapping
태백벼/가야벼 F3 집단 180 계통에 대해서 벼멸구저항성 유묘검정을 수행한 결과, F3 계통들의 계통별 저항성 지수 평균값은
Fig. 5과 같은 분포를 보였다. F3 계통들의 계통별 저항성 지수 평균값과 개발된 CAPS 마커 및
Bph3IND 마커와
Bph26IND 마커로 작성한 유전지도를 종합하여 저항성 QTL을 분석하였다. 그 결과 4번 염색체 0 - 6.6 cM 영역에서 LOD 값이 7.22인 QTL인
qBPH4가 발견되었고, 12번 염색체 40.8 – 54.4 cM 영역에서 LOD 값이 6.64인 QTL인
qBPH12가 발견되었다(
Table 4). 확률 수준 0.05에서 1,000번의 permutation을 함으로서 결정된 LOD threshold 값은 3.9이었다.
qBPH4 영역은
Bph3IND 마커(5.7 cM)을 포함하고 있었고,
qBPH12 영역은
BPH26IND 마커(48.4 cM)을 각각 포함하고 있었다. 따라서,
qBPH4는
Bph3 유전자이며,
qBPH12는
BPH26 유전자로 판단되며, 이 결과는 가야벼의 벼멸구 저항성 유전자가
Bph3와
BPH26임을 뒷받침해주고 있다고 판단된다.
Fig. 5Distribution of BPH resistance scores of the 180 F3 families.
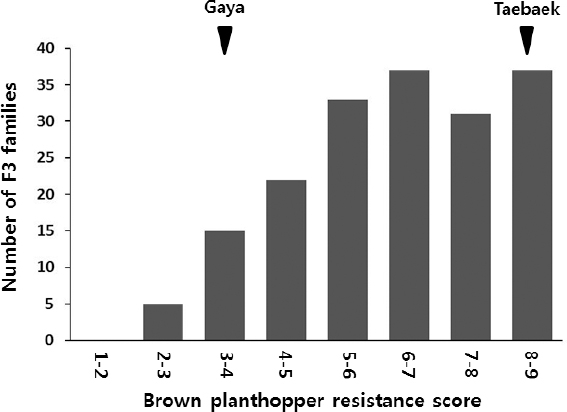
Table 4Identification of QTLs for BPH resistance
Table 4
|
QTL name |
chromosome |
location (cM) |
QTL interval* (cM) |
LOD |
additive effect |
dominance effect |
R2
|
|
qBPH4
|
4 |
0 |
0-6.6 |
7.22 |
0.94 |
-0.20 |
0.138 |
|
qBPH12
|
12 |
51.4 |
40.8-54.4 |
6.64 |
0.87 |
0.55 |
0.131 |
적요
통일형 벼품종인 가야벼는 우리나라에서 수집된 다양한 벼멸구 집단들에 대해서 저항성을 가진 것으로 알려져 있다. 가야벼 벼멸구 저항성의 유전분석을 위해서, 가야벼와 벼멸구 감수성 통일형 품종인 태백벼를 교배하여 F2 및 F3 집단을 작성하였다. 그리고, 선행연구에서 탐색된 가야벼와 태백벼간 약 28만개의 단일염기서열변이(SNPs)들 중 제한효소 인식부위에 있는 SNP들을 대상으로 99 개의 CAPS (Cleaved Amplified Polymorphic Sequences) 마커를 개발하였다. 이들 마커들로 태백벼/가야벼 F2 집단 180 개체들의 유전자형을 분석하여 유전지도를 작성하였다. 가야벼의 유전체재해석(resequencing) 염기서열 데이터와 기존에 알려진 벼멸구 저항성 유전자들(Bph3, BPH9, Bph14, BPH18, BPH26)의 염기서열을 비교한 결과, 가야벼가 Bph3과 BPH26 유전자들을 가지고 있는 것으로 나타났다. 가야벼의 Bph3과 BPH26 유전자 영역을 PCR 증폭하여 Sanger sequencing 방법으로 염기서열을 분석한 결과, Bph3 및 BPH26 유전자 염기서열과 완전히 일치함을 확인하였다. 이 두 유전자 영역에서 삽입/결실(InDel) 마커인 Bph3IND와 BPH26IND를 개발하였고, 이들 마커들은 가야벼의 저항성 유전자들을 고품질 자포니카 벼 품종들에 도입하는 육종 프로그램에 선발마커로서 활용될 수 있음을 확인하였다. 태백벼/가야벼 F3 집단 180 계통들을 대상으로 벼멸구저항성 유묘검정을 수행하여 QTL 분석을 한 결과, 4번 염색체와 12번 염색체에서 주동 QTL 두 개가 발견되었다. 이 두 QTL들의 영역은 각각 Bph3과 BPH26 유전자를 포함하고 있었는데, 이는 가야벼가 Bph3과 BPH26 유전자를 가지고 있음을 뒷받침해 주었다. 이 결과는 우리나라에서 고품질 벼멸구저항성 자포니카 벼 품종 육성을 촉진하는데 유용하게 활용될 수 있을 것이다.
사사
본 논문은 농촌진흥청 연구사업(과제번호: PJ01104201)의 지원에 의해 이루어진 것임.
References
- Basten CJ, Weir BS, Zeng ZB. 1996. QTL Cartographer: A reference manual and tutorial for QTL mapping. Department of Statistics, North Carolina State University. Raleigh, NC.
- Cantindig JLA, Arida GS, Baehaki SE, Bentur JS, Cuong LQ, Norowi M, Rattanakam W, Sriratanasak W, Xia J, Lu Z. 2009. Situation of planthoppers in Asia. Heong KL, Hardy B, editors. Planthoppers: new threats to the sustainability of intensive rice production systems in Asia. International Rice Research Institute. Los Baños, Philippines: pp. 191-220.
- Du B, Zhang W, Liu B, Hu J, Wei Z, Shi Z, He R, Zhu L, Chen R, Han B, He G. 2009. Identification and characterization of Bph14 a gene conferring resistance to brown planthopper in rice. Proc Natl Acad Sci U S A 106: 22163-22168.
- Fujita D, Kohli A, Horgan FG. 2013. Rice Resistance to Planthoppers and Leafhoppers. Critical Reviews in Plant Sciences 32: 162-191.
- Hibino H. 1996. Biology and epidemiology of rice viruses. Annu Review of Entomology 34: 249-274.
- Hu J, Cheng MX, Gao GJ, Zhang QL, Xiao JH, He Y. 2013. Pyramiding and evaluation of three dominant brown planthopper resistance genes in the elite indica rice 9311 and its hybrids. Pest Management Science 69: 802-808.
- Hu J, Xiao C, He Y. 2016. Recent progress on the genetics and molecular breeding of brown planthopper resistance in rice. Rice (N Y) 9: 30
- Huang Z, He G, Shu L, Li X, Zhang Q. 2001. Identification and mapping of two brown planthopper resistance genes in rice. Theor Appl Genet 102: 929-934.
- Ji H, Ahn E, Seo BY, Kang HJ, Choi I, Kim KH. 2016. Genome-wide detection of SNPs between two Korean Tongil-type rice varieties. Korean J Breed Sci 48: 460-469.
- Ji H, Kim SR, Kim YH, Suh JP, Park HM, Sreenivasulu N, Misra G, Kim SM, Hechanova SL, Kim H, Lee GS, Yoon UH, Kim TH, Lim H, Suh SC, Yang J, An G, Jena KK. 2016. Map-based cloning and characterization of the BPH18 gene from wild rice conferring resistance to brown planthopper (BPH) insect pest. Sci Rep 6: 34376
- Kim DH, Lee GH, So JD, Yuh JH, Kim JH. 1991. Resistance aspects on different growth stages and field resistance of rice varieties to brown planthopper Nilaparvata lugens STAL (Homoptera : Delphacidae). Research Report of RDA 33: 33-39.
- Kim YH, Lee JO, Choi KM, Cho SY. 1989. Varietal resistance of Korea rice cultivars to brown planthopper and whitebacked planthopper and their resistance sources. Research Report of RDA 31: 7-13.
- Lee JO, Goh HG, Kim YH. 1985. Study on brown planthopper biotypes. 2. Resistance mechanism of different rice varieties to Korean brown planthopper biotypes. Research Report of RDA 27: 73-78.
- Lee JO, Goh HG, Kim HS, Choi KM. 1986. Study on brown planthopper biotypes. 3. Plant damage of different rice cultivars to combination of biotypes. Research Report of RDA 28: 52-55.
- Liu Y, Wu H, Chen H, Liu Y, He J, Kang H, Sun Z, Pan G, Wang Q, Hu J, Zhou F, Zhou K, Zheng X, Ren Y, Chen L, Wang Y, Zhao Z, Lin Q, Wu F, Zhang X, Guo X, Cheng X, Jiang L, Wu C, Wang H, Wan J. 2014. A gene cluster encoding lectin receptor kinases confers broad-spectrum and durable insect resistance in rice. Nat Biotechnol 33: 301-305.
- Lorieux M. 2012. MapDisto: fast and efficient computation of genetic linkage maps. Mol Breeding 30: 1231-1235.
- Myint KK, Fujita D, Matsumura M, Sonoda T, Yoshimura A, Yasui H. 2012. Mapping and pyramiding of two major genes for resistance to the brown planthopper (Nilaparvata lugens [Stal]) in the rice cultivar ADR52. Theor Appl Genet 124: 495-504.
- Otuka A. 2009. Migration of rice planthoppers and simulation techniques. Heong KL, Hardy B, editors. Planthoppers: new threats to the sustainability of intensive rice production systems in Asia. International Rice Research Institute. Los Baños, Philippines: pp. 221-230.
- Park YD, Song YH. 1988. Preference, development and fecundity of the brown planthopper (Nilaparvata lugens Stal) biotypes fed on different cultivars of rice with various resistance gene. Korean Journal of Applied Entomology 27: 87-93.
- Qiu Y, Guo J, Jing S, Zhu L, He G. 2012. Development and characterization of japonica rice lines carrying the brown planthopper-resistance genes BPH12 and BPH6. Theor Appl Genet 124: 485-494.
- Seo BY, Jung JK, Choi BR, Park HM, Lee BH. 2009. Resistance-breaking ability and feeding behavior of the brown planthopper Nilaparvata lugens recently collected in Korea. Heong KL, Hardy B, editors. Planthoppers: new threats to the sustainability of intensive rice production systems in Asia. International Rice Research Institute. Los Baños, Philippines: pp. 303-314.
- Seo BY, Kwon YH, Jung JK, Kim GH. 2009. Electrical penetration graphic waveforms in relation to the actual positions of the stylet tips of Nilaparvata lugens in rice tissue. Asia-Pacific Entomology 12: 89-95.
- Sogawa K. 1982. The rice brown planthopper: feeding physiology and host plant interactions. Annual Review of Entomology 27: 49-73.
- Tamura Y, Hattori M, Yoshioka H, Yoshioka M, Takahashi A, Wu J, Sentoku N, Yasui H. 2014. Map-based cloning and characterization of a brown planthopper resistance gene BPH26 from Oryza sativa L. ssp. indica cultivar ADR52. Sci Rep 4: 5872
- Uhm KB, Park JS, Lee YI, Choi KM, Lee MH, Lee JO. 1988. Relationship between some weather conditions and immigration of the brown planthopper Nilaparvata lugens Stål. Koream Journal of Applied Entomology 27: 200-210.
- Wilson MR, Claridge MF. 1991. Handbook for the identification of leafhoppers and planthoppers of rice. CAB International. Wallingford, UK.
- Zhao Y, Huang J, Wang Z, Jing S, Wang Y, Ouyang Y, Cai B, Xin XF, Liu X, Zhang C, Pan Y, Ma R, Li Q, Jiang W, Zeng Y, Shangguan X, Wang H, Du B, Zhu L, Xu X, Feng YQ, He SY, Chen R, Zhang Q, He G. 2016. Allelic diversity in an NLR gene BPH9 enables rice to combat planthopper variation. Proc Natl Acad Sci U S A 113: 12850-12855.