

National Institute of Crop Science, RDA, Wanju, 55365, Korea
© Korean Society of Breeding Science. All rights reserved.
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
| Gene | Chromosome | Marker | Sequence (5'-3') | Product size (bp) | Reference |
|---|---|---|---|---|---|
|
|
|||||
| GW2 | 2 | GW2-HpaI | F: CCAATAAAGATGTCCATTCTGTTA | 23+151/174 | Yan et al. 2009 |
| R: GCTCTTCCTGTAACACATATTATG | (dCAPS/Hpa I) | Yan et al. 2011 | |||
| GS3 | 3 | GS3-PstI | F: TATTTATTGGCTTGATTTCCTGTG | 294+218/512 | Yan et al. 2009 |
| R: GCTGGTTTTTTACTTTCATTTGCC | (CAPS/Pst I) | Yan et al. 2011 | |||
| qSW5 | 5 | N1212del | F: CGTCTTGCAACCAACGCCGATGTTATAC | 759/1021/1971 | Shomura et al. 2008 |
| R: GAGCGTGTGTAGGGAAGGAGCTGCATGA | (Indel) | Yan et al. 2011 | |||
| Cross combination | Anther cultured (no.) | Cali induced (no. and %) | Plant regeneration (no. and %)z | Green plants (no. and %)y | Albino (no. and %)x | Doubled haploid (DH) lines (no.)w | Selected DH lines (no.)v |
|---|---|---|---|---|---|---|---|
|
|
|||||||
| Deuraechan/Jizi1560 | 10,700 | 6,280(58.7) | 1241(19.8) | 146(11.8) | 1095(88.2) | 99 | 88 |
| Deuraechan/Jizi1581 | 11,600 | 2,780(24.0) | 577(20.8) | 110(18.8) | 467(80.9) | 105 | 70 |
| Boramchan/Jizi1560 | 11,800 | 4,320(36.6) | 817(18.9) | 112(13.7) | 705(86.3) | 72 | 62 |
| Boramchan/Jizi1581 | 6,300 | 4,380(69.5) | 1184(27.0) | 228(19.3) | 956(80.7) | 84 | 70 |
| Total | 360 | 290 | |||||
zPercent plant regeneration is the number of plants regenerated over number of cali plated
yPercent green is the number of green plants regenerated over total plants regenerated
xPercent albino is the number of albino plants regenerated over total plants regenerated
wDouble haploid lines are harvested plants having fertile grain
vSelected DH lines are used in this study
| Trait | GW | GT | RLW | TGW |
|---|---|---|---|---|
|
|
||||
| GLz | 0.330**y | 0.285** | 0.638** | 0.789** |
| GW | 0.817** | -0.510** | 0.658** | |
| GT | -0.399** | 0.655** | ||
| RLW | 0.174** | |||
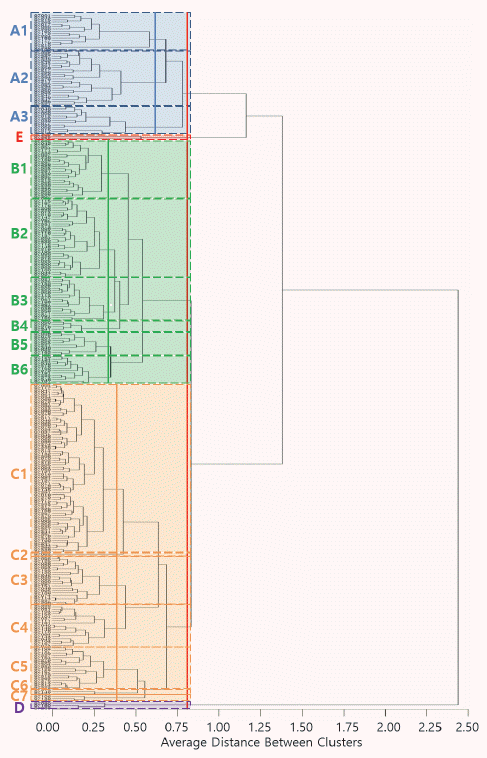
| Group | n | Rough rice | HD (DAS) | CL (cm) | PL (cm) | PN | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||||
| GLz (mm) | GW (mm) | GT (mm) | RLW | TGW (g) | |||||||
|
|
|||||||||||
| A | A1 | 15 | 10.42by | 4.08c | 2.54cd | 2.56c | 46.0b | 96bc | 84abc | 23bcde | 9abcd |
| A2 | 25 | 9.76c | 3.62efg | 2.37fgh | 2.70b | 37.9def | 97bc | 80bc | 24abc | 8bcd | |
| A3 | 13 | 9.69c | 4.46b | 2.68b | 2.18f | 43.3bc | 104bc | 80bc | 23cde | 8cd | |
| Mean | 53 | 9.93Cx | 3.96B | 2.49B | 2.53B | 41.5B | 98B | 81B | 24AB | 8B | |
| C.V.(%) | 53 | 3.9 | 9.8 | 6.4 | 9.5 | 11.5 | 11.2 | 13.8 | 10.1 | 24.7 | |
|
|
|||||||||||
| B | B1 | 24 | 8.70f | 3.78ef | 2.40efg | 2.31de | 35.0ef | 96bc | 83abc | 23cde | 9abcd |
| B2 | 36 | 8.79ef | 4.10c | 2.53cd | 2.15f | 37.7def | 99bc | 79bc | 22cde | 8abcd | |
| B3 | 19 | 8.44g | 4.21c | 2.60bc | 2.01g | 37.1def | 101bc | 77bc | 23cde | 8abcd | |
| B4 | 5 | 8.97e | 4.52b | 2.61bc | 1.98gh | 38.3de | 105bc | 72c | 22cde | 8bcd | |
| B5 | 10 | 9.32d | 4.15c | 2.50cde | 2.25ef | 40.1cd | 109b | 89ab | 26a | 7d | |
| B6 | 13 | 9.23d | 3.81de | 2.37fgh | 2.43d | 35.9ef | 95bc | 76bc | 23cde | 9abcd | |
| Mean | 107 | 8.82D | 4.04B | 2.49B | 2.20C | 37.0C | 99B | 80B | 23B | 8B | |
| C.V.(%) | 107 | 3.6 | 5.9 | 4.8 | 7.3 | 9.3 | 13.1 | 15.3 | 10.5 | 21.2 | |
|
|
|||||||||||
| C | C1 | 73 | 8.10h | 4.01cd | 2.53cd | 2.03g | 34.2f | 97bc | 75bc | 22cde | 9abcd |
| C2 | 1 | 7.59i | 4.05c | 2.42def | 1.88h | 30.2g | 106b | 80bc | 20e | 9abcd | |
| C3 | 22 | 7.77i | 3.63efg | 2.34fgh | 2.15f | 29.3g | 97bc | 75bc | 22cde | 9abcd | |
| C4 | 18 | 7.19j | 3.59fg | 2.34fgh | 2.01g | 27.0g | 100bc | 76bc | 21de | 10abc | |
| C5 | 19 | 8.22gh | 3.44gh | 2.29ghi | 2.40d | 29.8g | 105bc | 83abc | 23bcde | 9abcd | |
| C6 | 2 | 7.68i | 3.16ij | 2.09j | 2.43d | 23.5h | 98cb | 72c | 21e | 10abc | |
| C7 | 3 | 8.80ef | 3.30hi | 2.20i | 2.67bc | 29.9g | 109b | 79bc | 22cde | 9abcd | |
| Mean | 138 | 7.95E | 3.79B | 2.43B | 2.11C | 31.6D | 99B | 76B | 22B | 9AB | |
| C.V.(%) | 138 | 5.0 | 7.9 | 5.8 | 8.5 | 11.7 | 10.1 | 11.8 | 8.7 | 17.8 | |
|
|
|||||||||||
| D | D | 3 | 11.56aA | 5.26aA | 2.97aA | 2.20efC | 55.3aA | 89cB | 83abcAB | 24abcdAB | 8cdB |
| C.V.(%) | 3 | 1.5 | 3.6 | 1.7 | 2.7 | 2.9 | 0.0 | 3.4 | 8.5 | 13.2 | |
|
|
|||||||||||
| E | E | 2 | 10.60bB | 3.07jC | 2.27hiC | 3.45aA | 35.0eCD | 125aA | 95aA | 26abA | 11aA |
| C.V.(%) | 2 | 4.8 | 1.6 | 2.6 | 3.2 | 1.2 | 0.0 | 3.3 | 0.8 | 41.7 | |
|
|
|||||||||||
| Total | Mean | 303 | 8.66 | 3.91 | 2.47 | 2.23 | 35.5 | 99 | 78 | 23 | 9 |
| C.V.(%) | 303 | 10.2 | 9.0 | 6.1 | 11.7 | 16.0 | 12.1 | 13.8 | 10.2 | 20.7 | |
zGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight, HD: heading date, DAS: days after seeding, CL: culm length, PL: panicle length, PN: number of panicles per hill
yMeans with the same letters in a column are not significantly different at p<0.05 (ANOVA followed by DMRT)
xCapital letters indicate statistic difference among main groups defined by culstering analysis
| Group | n | Rough rice | Brown rice | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
||||||||||||
| GLz (mm) | GW (mm) | GT (mm) | RLW | TGW (g) | GL (mm) | GW (mm) | GT (mm) | RLW | TGW (g) | |||
|
|
||||||||||||
| A | A1 | 6 | 10.43by | 4.19cd | 2.57bc | 2.49bc | 47.1b | 7.64b | 3.42bcde | 2.31abcd | 2.24bc | 39.8b |
| A2 | 10 | 9.49c | 3.62ef | 2.37def | 2.63b | 37.7de | 7.04c | 3.09fgh | 2.14cdef | 2.29b | 32.9de | |
| A3 | 8 | 9.53c | 4.53bc | 2.70b | 2.12efg | 45.2bc | 6.81cd | 3.60ab | 2.41a | 1.90def | 37.5bc | |
| Mean | 24 | 9.74Bx | 4.07B | 2.53B | 2.43B | 42.6B | 7.12B | 3.34A | 2.27AB | 2.15BC | 36.1B | |
| C.V.(%) | 24 | 4.7 | 11.6 | 7.0 | 10.9 | 11.6 | 5.5 | 8.6 | 6.5 | 10.4 | 9.6 | |
|
|
||||||||||||
| B | B1 | 6 | 8.88d | 3.87de | 2.37ef | 2.30cde | 35.2efg | 6.39de | 3.25cdef | 2.18bcdef | 1.97de | 30.6def |
| B2 | 13 | 8.87d | 4.18cd | 2.56bcd | 2.13defg | 39.7de | 6.32ef | 3.49abcd | 2.31abc | 1.81efg | 33.1de | |
| B3 | 7 | 8.31e | 4.23bcd | 2.54bcde | 1.97fg | 37.1de | 5.96fgh | 3.57ab | 2.35ab | 1.67gh | 31.5de | |
| B4 | 4 | 8.79d | 4.58b | 2.70b | 1.93g | 41.5cd | 6.15efg | 3.72a | 2.41a | 1.66gh | 34.1cd | |
| B5 | 4 | 9.38c | 4.24bcd | 2.51bcde | 2.22cdef | 40.9d | 6.81cd | 3.51abc | 2.22bcde | 1.95de | 34.0cd | |
| B6 | 4 | 9.30c | 3.94de | 2.40cdef | 2.63b | 37.5de | 6.77cd | 3.23def | 2.17bcdef | 2.10bcd | 31.5de | |
| Mean | 38 | 8.86C | 4.16B | 2.52B | 2.17B | 38.6BC | 6.34C | 3.47A | 2.28AB | 1.84CD | 32.4BC | |
| C.V.(%) | 38 | 4.8 | 6.3 | 5.6 | 12.5 | 8.5 | 6.0 | 5.5 | 5.9 | 9.8 | 7.0 | |
|
|
||||||||||||
| C | C1 | 9 | 8.02ef | 4.17cd | 2.58bc | 1.93g | 35.5ef | 5.64hij | 3.49abcd | 2.35ab | 1.62gh | 29.5ef |
| C2 | 1 | 7.49g | 3.94de | 2.36ef | 1.90g | 29.9h | 5.24jk | 3.34bcdef | 2.13cdef | 1.57h | 24.9ghi | |
| C3 | 5 | 7.58g | 3.64ef | 2.36ef | 2.09efg | 30.1h | 5.46ijk | 3.20efg | 2.12def | 1.71fgh | 25.0ghi | |
| C4 | 5 | 7.25g | 3.66ef | 2.34ef | 1.99fg | 28.0hi | 5.18k | 3.08fgh | 2.13cdef | 1.69gh | 22.6hi | |
| C5 | 6 | 8.06ef | 3.35f | 2.26fg | 2.41bcd | 28.9hi | 5.80ghi | 2.83hi | 2.03ef | 2.05cd | 24.0hi | |
| C6 | 2 | 7.66fg | 3.50f | 2.24gf | 2.22cdef | 24.9i | 5.49ijk | 2.89hi | 1.99f | 1.93de | 21.8i | |
| C7 | 3 | 8.37e | 3.48f | 2.29fg | 2.42bc | 31.1gh | 6.20efg | 2.96gh | 2.07ef | 2.11bcd | 25.9gh | |
| Mean | 31 | 7.83D | 3.73B | 2.39BC | 2.12B | 30.8D | 5.60D | 3.16A | 2.16CB | 1.79D | 25.6D | |
| C.V.(%) | 31 | 5.3 | 9.7 | 6.7 | 10.6 | 13.1 | 6.2 | 9.1 | 7.6 | 12.1 | 13.6 | |
|
|
||||||||||||
| D | D | 1 | 11.52aA | 5.41aA | 2.97aA | 2.13defgB | 56.6aA | 8.17aA | 3.57abA | 2.45aA | 2.29bB | 48.1aA |
|
|
||||||||||||
| E | E | 2 | 9.65cB | 3.00gC | 2.14gC | 3.22aA | 32.4fghCD | 7.52bB | 2.69iB | 1.99fC | 2.81aA | 27.9fgCD |
|
|
||||||||||||
| Total | Mean | 96 | 8.79 | 4.00 | 2.47 | 2.24 | 37.1 | 6.34 | 3.32 | 2.23 | 1.93 | 31.2 |
| C.V.(%) | 96 | 10.2 | 11.2 | 7.3 | 13.9 | 17.2 | 11.5 | 8.9 | 7.2 | 14.5 | 17.3 | |
zGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
yMeans with the same letters in a column are not significantly different at p<0.05 (ANOVA followed by DMRT)
xCapital letters indicate statistic difference among main groups defined by culstering analysis
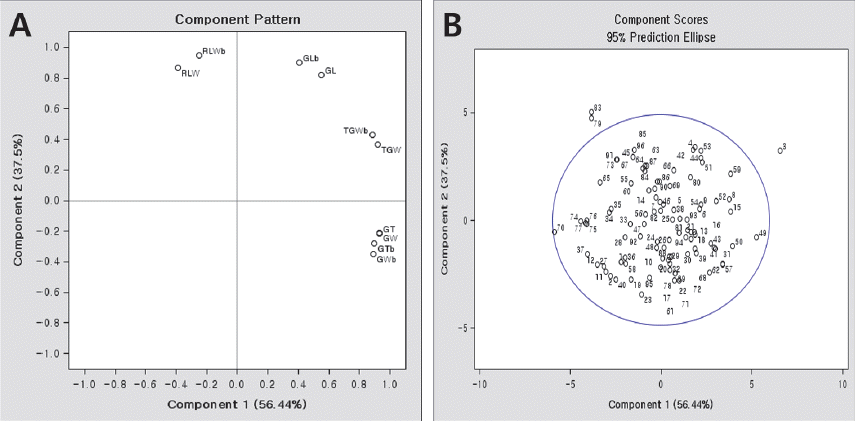
| Trait | Eigenvector | Correlation coefficient | ||||
|---|---|---|---|---|---|---|
|
|
||||||
| Prin1y | Prin2 | Prin3 | Prin1 | Prin2 | Prin3 | |
|
|
||||||
| GLz | 0.232 | 0.425 | -0.171 | 0.551**x | 0.822** | -0.089ns |
| GW | 0.392 | -0.109 | -0.411 | 0.931** | -0.212* | -0.213* |
| GT | 0.393 | -0.112 | 0.422 | 0.933** | -0.218* | 0.218* |
| RLW | -0.165 | 0.447 | 0.280 | -0.391** | 0.865** | 0.145ns |
| TGW | 0.386 | 0.189 | -0.028 | 0.918** | 0.365** | -0.014ns |
| GLbw | 0.170 | 0.466 | -0.180 | 0.405** | 0.902** | -0.093ns |
| GWb | 0.375 | -0.179 | -0.320 | 0.890** | -0.347** | -0.166ns |
| GTb | 0.376 | -0.143 | 0.627 | 0.894** | -0.277** | 0.325** |
| RLWb | -0.106 | 0.491 | 0.122 | -0.253* | 0.950** | 0.063ns |
| TGWb | 0.373 | 0.223 | 0.052 | 0.887** | 0.431** | 0.027ns |
|
|
||||||
| Eigen value | 5.64 | 3.75 | 0.27 | |||
| Contribution (%) | 56.4 | 37.5 | 2.7 | |||
| Cumulative contribution (%) | 56.4 | 93.9 | 96.6 | |||
zGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
yPrin1, Prin2, and Prin3 indicate the first, second, and third component, respectively
xns, *, and ** mean no significant, significant at p<0.05, and p<0.01, respectively
wb means brown rice
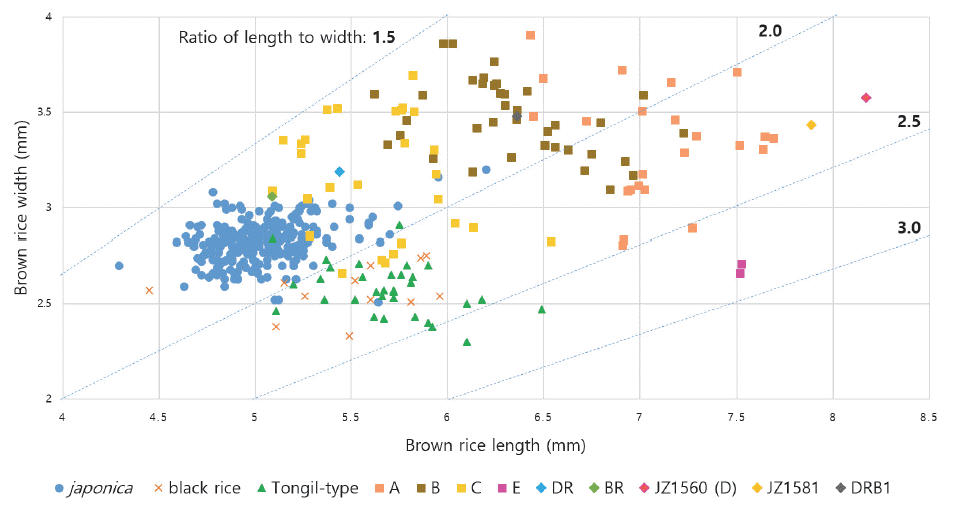
| Group | n | GLz (mm) | GW (mm) | GT (mm) | RLW | TGW (g) |
|---|---|---|---|---|---|---|
|
|
||||||
| Deuraechan | 1 | 5.44 | 3.19 | 2.12 | 1.71 | 24.7 |
| Boramchan | 1 | 5.09 | 3.06 | 2.13 | 1.67 | 22.9 |
| Jizi1560 | 1 | 8.17 | 3.57 | 2.45 | 2.29 | 48.1 |
| Jizi1581 | 1 | 7.89 | 3.43 | 2.29 | 2.30 | 38.0 |
| Daeripbyeo 1 | 1 | 6.36 | 3.48 | 2.25 | 1.83 | 34.6 |
|
|
||||||
| DH lines | 91 | 6.34±0.68ay | 3.32±0.30a | 2.23±0.16a | 1.93±0.28c | 31.1±5.1a |
| (5.09-7.69) | (2.66-3.91) | (1.83-2.63) | (1.53-2.83) | (18.6-41.5) | ||
| japonica | 264 | 5.03±0.23dx | 2.82±0.11b | 1.94±0.13b | 1.79±0.09d | 22.0±2.0b |
| (4.29-6.20) | (2.51-3.20) | (1.23-2.30) | (1.55-2.02) | (14.1-34.8) | ||
| black rice | 13 | 5.34±0.63c | 2.57±0.12c | 1.78±0.11c | 2.08±0.25b | 19.5±2.9c |
| (3.76-5.96) | (2.33-2.75) | (1.57-1.97) | (1.46-2.36) | (14.8-23.5) | ||
| Tongil-type | 32 | 5.69±0.30b | 2.57±0.13c | 1.83±0.13c | 2.22±0.19a | 22.5±2.1b |
| (5.09-6.49) | (2.30-2.91) | (1.60-2.14) | (1.79-2.65) | (18.9-28.2) | ||
zGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
yMeans with the same letters in a column are not significantly different at p<0.05 (ANOVA followed by DMRT)
xThe data of japonica, black rice, and Tongil-type cultivars were used the value on registration of national variety and in the website of 'Nongsaro' (http://www.nongsaro.go.kr/)
| Group | n | HDz (DAS) | CL (cm) | PL (cm) | PN | NS | RRG (%) | Yield (g) | |
|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||
| A | A1 | 6 | 95bcy | 80ab | 24abc | 11abc | 68de | 66.6abcd | 468cdef |
| A2 | 10 | 94bc | 80ab | 26a | 9bc | 136ab | 52.7de | 494bcdef | |
| A3 | 8 | 102bc | 84ab | 23abc | 9abc | 88cde | 39.9e | 405ef | |
| Mean | 24 | 97Bx | 82B | 24A | 9A | 103AB | 51.9A | 458AB | |
| C.V.(%) | 24 | 10.5 | 13.9 | 8.2 | 19.6 | 38.2 | 36.2 | 16.7 | |
|
|
|||||||||
| B | B1 | 6 | 97bc | 79b | 24abc | 11abc | 95bcde | 55.7cde | 483bcdef |
| B2 | 13 | 102bc | 80ab | 23abc | 10abc | 94bcde | 68.2abcd | 511abcdef | |
| B3 | 7 | 106bc | 81ab | 23abc | 8c | 125abc | 59.8bcde | 527abcde | |
| B4 | 4 | 107bc | 73b | 22bc | 10abc | 93bcde | 53.4de | 453def | |
| B5 | 4 | 109abc | 90ab | 24abc | 8c | 112abc | 54.1de | 446def | |
| B6 | 4 | 98bc | 82ab | 25ab | 11abc | 105abcd | 56.5cde | 544abcde | |
| Mean | 38 | 103B | 81B | 23A | 10A | 103AB | 60.4A | 500AB | |
| C.V.(%) | 38 | 13.2 | 14.4 | 8.3 | 19.5 | 24.2 | 26.2 | 17.9 | |
|
|
|||||||||
| C | C1 | 9 | 99bc | 75b | 23abc | 10abc | 118abc | 64.7abcde | 523abcdef |
| C2 | 1 | 109abc | 83ab | 21c | 12a | 123abc | 84.6ab | 632abc | |
| C3 | 5 | 109abc | 76b | 22bc | 12ab | 118abc | 77.5abcd | 554abcde | |
| C4 | 5 | 104bc | 76b | 21c | 12ab | 135ab | 81.5abc | 641ab | |
| C5 | 6 | 115ab | 83ab | 21c | 10abc | 125abc | 86.9a | 596abcd | |
| C6 | 2 | 104bc | 73b | 22bc | 11abc | 150a | 69.1abcd | 666a | |
| C7 | 3 | 110abc | 81ab | 23abc | 10abc | 122abc | 78.8abcd | 556abcde | |
| Mean | 31 | 106B | 78B | 22A | 11A | 125A | 76.1A | 577A | |
| C.V.(%) | 31 | 10.3 | 8.2 | 8.1 | 18.0 | 18.5 | 15.4 | 17.8 | |
|
|
|||||||||
| D | D | 1 | 92cB | 91abAB | 24abcA | 10abcA | 55eB | 7.1fB | 362fB |
|
|
|||||||||
| E | E | 2 | 128aA | 98aA | 24abcA | 10abcA | 123abcA | 73.8abcdA | 542abcdeA |
|
|
|||||||||
| Total | Mean | 96 | 103 | 80 | 23 | 10 | 110 | 63.0 | 513 |
| C.V.(%) | 96 | 12.4 | 12.9 | 8.8 | 18.4 | 27.7 | 29.7 | 19.9 | |
zHD: heading date, DAS: days after seeding, CL: culm length, PL: panicle length, PN: number of panicles per hill, NS: number of spikelets per panicle, RRG: ratio of ripened grain
yMeans with the same letters in a column are not significantly different at p<0.05 (ANOVA followed by DMRT)
xCapital letters indicate statistic difference among main groups defined by culstering analysis
| Traitz | GLy | GW | GT | RLW | TGW | HD | CL | PL | PN | NS | RRG | Yield |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (x1, 6) | (x2, 7) | (x3, 8) | (x4, 9) | (x5, 10) | (x11) | (x12) | (x13) | (x14) | (x15) | (x16) | (x17) | |
|
|
||||||||||||
| x1, 6 | 0.364** | 0.322** | 0.478** | 0.801** | -0.255* | 0.237* | 0.475** | -0.168ns | -0.427** | -0.508** | -0.476** | |
| x2, 7 | 0.066nsx | 0.887** | -0.576** | 0.768** | -0.310** | -0.127ns | 0.043ns | -0.230* | -0.463** | -0.561** | -0.491** | |
| x3, 8 | 0.088ns | 0.850** | -0.534** | 0.770** | -0.190ns | -0.068ns | 0.038ns | -0.249* | -0.431** | -0.417** | -0.427** | |
| x4, 8 | 0.759** | -0.591** | -0.467** | -0.043ns | 0.075ns | 0.255* | 0.396** | 0.037ns | 0.048ns | 0.080ns | 0.012ns | |
| x5, 10 | 0.741** | 0.639** | 0.680** | 0.177ns | -0.270** | 0.103ns | 0.292** | -0.213* | -0.576** | -0.498** | -0.554** | |
| x11 | -0.220* | -0.208* | -0.166ns | -0.024ns | -0.276** | 0.525** | -0.257* | -0.007ns | 0.290** | 0.440** | 0.434** | |
| x12 | 0.289** | -0.133ns | -0.090ns | 0.324** | 0.106 | 0.525** | 0.141ns | -0.192ns | 0.108ns | 0.115ns | 0.209* | |
| x13 | 0.477** | 0.057ns | 0.033ns | 0.355** | 0.366** | -0.257* | 0.141ns | -0.425** | 0.132ns | -0.324** | -0.274** | |
| x14 | -0.150ns | -0.269** | -0.285** | 0.033ns | -0.243* | -0.007ns | -0.192ns | -0.425** | -0.451** | 0.462** | 0.240* | |
| x15 | -0.345** | -0.381** | -0.359** | -0.022ns | -0.506** | 0.290** | 0.108ns | 0.132ns | -0.451** | 0.038ns | 0.507** | |
| x16 | -0.447** | -0.398** | -0.375** | -0.100ns | -0.522** | 0.440** | 0.115ns | -0.324** | 0.462** | 0.038ns | 0.575** | |
| x17 | -0.420** | -0.410** | -0.367** | -0.081ns | -0.529** | 0.434** | 0.209* | -0.274** | 0.240* | 0.507** | 0.575** | |
zUpper and lower diagonal indicate the value of rough and brown rice, respectively
yGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight, HD: heading date, CL: culm length, PL: panicle length, PN: number of panicles per hill, NS: number of spikelets per panicle, RRG: ratio of ripened grain
xns, *, and ** mean no significant, significant at p<0.05, and p<0.01, respectively
| Group | n | GW2 | GS3 | qSW5 | Remark | ||||
|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||
| gw2 | GW2 | gs3 | GS3 | qsw5 | qSW5 | ||||
|
|
|||||||||
| A1 | 6 | 3 | 3 | 6 | 0 | 5 | 1 | Jizi1581 (gw2gs3qSW5) | |
| A2 | 10 | 6 | 4 | 10 | 0 | 1 | 9 | ||
| A | A3 | 8 | 7 | 1 | 6 | 2 | 8 | 0 | |
| Total | 24 | 16 | 8 | 22 | 2 | 14 | 10 | ||
| (66.7)z | (33.3) | (91.7) | (8.3) | (58.3) | (41.7) | ||||
|
|
|||||||||
| B1 | 6 | 0 | 6 | 3 | 3 | 6 | 0 | Daeripbyeo 1 (gw2GS3qsw5) | |
| B2 | 13 | 9 | 4 | 3 | 10 | 11 | 2 | ||
| B3 | 7 | 6 | 1 | 2 | 5 | 7 | 0 | ||
| B | B4 | 4 | 4 | 0 | 1 | 3 | 4 | 0 | |
| B5 | 4 | 3 | 1 | 3 | 1 | 3 | 1 | ||
| B6 | 4 | 1 | 3 | 3 | 1 | 3 | 1 | ||
| Total | 38 | 23 | 15 | 15 | 23 | 34 | 4 | ||
| (60.5) | (39.5) | (39.5) | (60.5) | (89.5) | (10.5) | ||||
|
|
|||||||||
| C1 | 9 | 5 | 4 | 0 | 9 | 8 | 1 | ||
| C2 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | Deuraechan (GW2GS3qsw5) | |
| C3 | 5 | 0 | 5 | 0 | 5 | 5 | 0 | ||
| C4 | 5 | 0 | 5 | 0 | 5 | 4 | 1 | Boramchan (GW2GS3qsw5) | |
| C | C5 | 6 | 0 | 6 | 1 | 5 | 1 | 5 | |
| C6 | 2 | 1 | 1 | 0 | 2 | 1 | 1 | ||
| C7 | 3 | 0 | 3 | 1 | 2 | 1 | 2 | ||
| Total | 31 | 6 | 25 | 2 | 29 | 21 | 10 | ||
| (19.4) | (80.6) | (6.5) | (93.5) | (67.7) | (32.3) | ||||
|
|
|||||||||
| D | D | 1 | 1 | 0 | 1 | 0 | 1 | 0 | Jizi1560 (gw2gs3qsw5) |
| (100) | (0) | (100) | (0) | (100) | (0) | ||||
|
|
|||||||||
| E | E | 2 | 0 | 2 | 2 | 0 | 0 | 2 | DGS79, 83 (GW2gs3qSW5) |
| (0) | (100) | (100) | (0) | (0) | (100) | ||||
|
|
|||||||||
| Total | 96 | 46 | 50 | 42 | 54 | 70 | 26 | ||
| (47.9) | (52.1) | (43.8) | (56.3) | (72.9) | (27.1) | ||||
| Source of variance | df | Rough rice | Brown rice | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||||||||||||||||||||||||
| GLz | GW | GT | RLW | TGW | GL | GW | GT | RLW | TGW | ||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||
| MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | ||
|
|
|||||||||||||||||||||||||||||||
| GW2 | 1 | 2.13 | 5.72*y | 0.035 | 5.61 | 90.34** | 0.340 | 0.71 | 53.57** | 0.276 | 1.26 | 39.77** | 0.156 | 668.91 | 32.63** | 0.202 | 0.19 | 0.92ns | 0.005 | 2.72 | 123.16** | 0.372 | 0.54 | 53.22** | 0.268 | 0.83 | 47.10** | 0.131 | 431.83 | 30.77** | 0.182 |
| GS3 | 1 | 25.04 | 67.36** | 0.411 | 0.00 | 0.01ns | 0.000 | 0.01 | 0.50ns | 0.003 | 2.15 | 67.75** | 0.266 | 417.16 | 20.35** | 0.126 | 20.71 | 98.71** | 0.513 | 0.04 | 1.70ns | 0.005 | 0.01 | 0.55ns | 0.003 | 2.43 | 137.72** | 0.383 | 412.70 | 29.41** | 0.174 |
| qSW5 | 1 | 0.10 | 0.26ns | 0.002 | 5.02 | 80.83** | 0.304 | 0.64 | 48.78** | 0.251 | 1.54 | 48.57** | 0.191 | 387.05 | 18.88** | 0.117 | 0.21 | 0.99ns | 0.005 | 2.39 | 108.30** | 0.327 | 0.54 | 52.58** | 0.265 | 1.29 | 73.23** | 0.204 | 235.22 | 16.76** | 0.099 |
| GW2*GS3 | 1 | 0.73 | 1.97ns | 0.012 | 0.00 | 0.02ns | 0.000 | 0.02 | 1.49ns | 0.008 | 0.18 | 5.65* | 0.022 | 11.85 | 0.58ns | 0.004 | 0.62 | 2.97ns | 0.015 | 0.01 | 0.28ns | 0.001 | 0.04 | 3.76ns | 0.019 | 0.09 | 5.00* | 0.014 | 25.40 | 1.81ns | 0.011 |
| GW2*qSW5 | 1 | 0.12 | 0.33ns | 0.002 | 0.09 | 1.41ns | 0.005 | 0.00 | 0.02ns | 0.000 | 0.03 | 0.89ns | 0.003 | 12.64 | 0.62ns | 0.004 | 0.06 | 0.29ns | 0.002 | 0.19 | 8.70** | 0.026 | 0.00 | 0.02ns | 0.000 | 0.10 | 5.52* | 0.015 | 27.71 | 1.97ns | 0.012 |
| GS3*qSW5 | 1 | 0.01 | 0.02ns | 0.000 | 0.26 | 4.17* | 0.016 | 0.00 | 0.22ns | 0.001 | 0.10 | 3.03ns | 0.012 | 5.44 | 0.27ns | 0.002 | 0.08 | 0.40ns | 0.002 | 0.00 | 0.13ns | 0.000 | 0.00 | 0.01ns | 0.000 | 0.05 | 2.84ns | 0.008 | 0.04 | 0.00ns | 0.000 |
| GW2*GS3*qSW5 | 1 | 0.00 | 0.00 ns | 0.000 | 0.05 | 0.79 ns | 0.003 | 0.02 | 1.46 ns | 0.008 | 0.02 | 0.57 ns | 0.002 | 2.06 | 0.10 ns | 0.001 | 0.00 | 0.00 ns | 0.000 | 0.02 | 0.82 ns | 0.002 | 0.01 | 0.59 ns | 0.003 | 0.00 | 0.01 ns | 0.000 | 0.36 | 0.03 ns | 0.000 |
| Model | 7 | 4.02 | 10.81** | 0.462 | 1.57 | 25.37** | 0.669 | 0.20 | 15.15** | 0.546 | 0.75 | 23.75** | 0.654 | 215.01 | 10.49** | 0.455 | 3.13 | 14.90** | 0.542 | 0.77 | 34.73** | 0.734 | 0.16 | 15.82** | 0.557 | 0.68 | 38.77** | 0.755 | 161.89 | 11.54** | 0.479 |
| Error | 88 | 0.372 | 0.062 | 0.013 | 0.032 | 20.50 | 0.210 | 0.022 | 0.010 | 0.018 | 14.03 | ||||||||||||||||||||
| Gene combination | Genotype | n | Rough rice | Brown rice | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
||||||||||||
| GLz (mm) | GW (mm) | GT (mm) | RLW | TGW (g) | GL (mm) | GW (mm) | GT (mm) | RLW | TGW (g) | |||
|
|
||||||||||||
| Single gene | gw2 | 46 | 9.05±0.797**y | 4.27±0.373** | 2.58±0.159** | 2.14±0.240** | 40.6±5.31** | 6.46±0.656ns | 3.50±0.211** | 2.33±0.139** | 1.86±0.237* | 34.0±4.16** |
| GW2 | 50 | 8.55±0.921 | 3.73±0.341 | 2.38±0.136 | 2.33±0.340 | 33.9±5.61 | 6.23±0.774 | 3.15±0.260 | 2.15±0.127 | 1.99±0.303 | 28.7± 5.15 | |
|
|
||||||||||||
| gs3 | 42 | 9.51±0.633** | 4.01±0.434ns | 2.46±0.174ns | 2.43±0.339** | 40.2±5.65** | 6.98±0.494** | 3.34±0.281ns | 2.22±0.132ns | 2.14±0.255** | 34.1±4.46** | |
| GS3 | 54 | 8.23±0.621 | 3.97±0.421 | 2.48±0.185 | 2.09±0.180 | 34.8±5.97 | 5.84±0.431 | 3.29±0.308 | 2.25±0.181 | 1.76±0.166 | 29.0±4.99 | |
|
|
||||||||||||
| qsw5 | 70 | 8.76±0.921ns | 4.14±0.366** | 2.53±0.161** | 2.14±0.250** | 38.3±6.33** | 6.26±0.714ns | 3.43±0.213** | 2.29±0.141** | 1.83±0.218** | 32.1±5.25** | |
| qSW5 | 26 | 8.88±0.829 | 3.56±0.372 | 2.32± 0.125 | 2.51±0.298 | 34.0±5.57 | 6.55±0.729 | 3.02±0.276 | 2.09±0.117 | 2.19±0.269 | 28.9±5.15 | |
|
|
||||||||||||
| Three genes | gw2gs3qsw5 | 14 | 9.57±0.852ax | 4.51±0.296a | 2.64±0.133a | 2.13±0.160c | 44.4±5.45a | 6.84±0.600a | 3.57±0.127a | 2.35±0.070a | 1.92±0.185de | 36.6±4.87a |
| gw2gs3qSW5 | 9 | 9.56±0.433a | 3.85±0.274b | 2.39±0.069b | 2.50±0.192b | 39.4±2.82b | 7.08±0.363a | 3.25±0.164b | 2.16±0.088 | 2.18±0.121b | 34.0±1.55ab | |
| gw2GS3qsw5 | 20 | 8.53±0.507b | 4.33±0.305a | 2.65±0.137a | 1.98±0.095c | 39.2±5.08b | 5.98±0.384b | 3.59±0.191a | 2.40±0.132a | 1.67±0.095g | 32.7±3.76abc | |
| gw2GS3qSW5 | 3 | 8.55±0.475b | 4.03±0.220b | 2.49±0.122b | 2.13±0.214c | 36.2±3.67bc | 6.09±0.361b | 3.36±0.123b | 2.24±0.072b | 1.81±0.150ef | 30.3±2.75bcd | |
| GW2gs3qsw5 | 13 | 9.51±0.606a | 3.93±0.155b | 2.43±0.120b | 2.50±0.297b | 39.3±5.00b | 6.98±0.503a | 3.26±0.104b | 2.20±0.087b | 2.15±0.119bc | 33.9±4.31ab | |
| GW2gs3qSW5 | 6 | 9.32±0.393a | 3.24±0.189c | 2.25±0.102c | 2.89±0.258a | 33.4±2.10c | 7.11±0.391a | 2.79±0.085c | 2.04±0.066c | 2.56±0.203a | 29.0±1.98cd | |
| GW2GS3qsw5 | 23 | 8.03±0.680b | 3.88±0.242b | 2.43±0.106b | 2.08±0.141c | 33.2±4.53c | 5.74±0.492b | 3.31±0.161b | 2.20±0.112b | 1.74±0.126gf | 27.8±3.94d | |
| GW2GS3qSW5 | 8 | 7.90±0.393b | 3.32±0.122c | 2.22±0.069c | 2.38±0.097b | 27.7±3.00d | 5.72±0.279b | 2.80±0.092c | 1.99±0.077c | 2.05±0.082cd | 22.5±2.39e | |
zGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
yns, *, and ** mean no significant, significant at p<0.05, and p<0.01 by t-test, respectively
xMeans with the same letters in a column are not significantly difference at p<0.05 (ANOVA followed by DMRT)
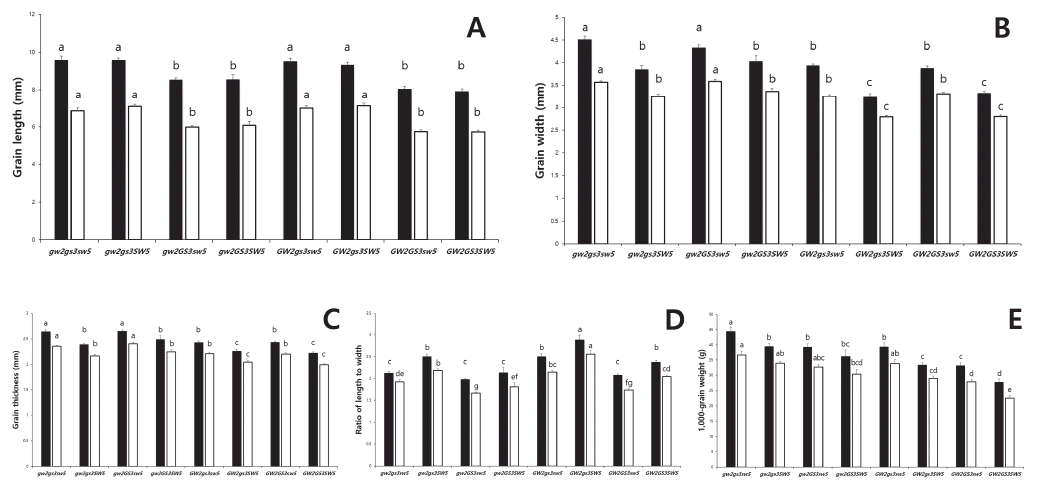
| Source of variance | df | HDz | CL | PL | PN | NS | RRG | Yield | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
||||||||||||||||||||||
| MS | MS | R 2 | MS | MS | R 2 | MS | MS | R 2 | MS | MS | R 2 | MS | MS | R 2 | MS | MS | R 2 | MS | MS | R 2 | ||
|
|
||||||||||||||||||||||
| GW2 | 1 | 456.17 | 3.03nsy | 0.030 | 69.24 | 0.66ns | 0.007 | 1.92 | 0.57ns | 0.005 | 71.89 | 32.69** | 0.251 | 68.69 | 0.09ns | 0.001 | 4710.60 | 23.28** | 0.161 | 25221.91 | 3.04ns | 0.027 |
| GS3 | 1 | 771.18 | 5.13* | 0.051 | 23.78 | 0.23ns | 0.002 | 65.81 | 19.50** | 0.163 | 4.44 | 2.02ns | 0.016 | 154.29 | 0.21ns | 0.002 | 5093.57 | 25.17** | 0.174 | 92956.71 | 11.19** | 0.099 |
| qSW5 | 1 | 149.60 | 1.00ns | 0.010 | 286.80 | 2.72ns | 0.028 | 0.29 | 0.09ns | 0.001 | 11.05 | 5.02* | 0.039 | 10612.05 | 14.16** | 0.128 | 993.58 | 4.91* | 0.034 | 59012.53 | 7.11** | 0.063 |
| GW2*GS3 | 1 | 111.21 | 0.74ns | 0.007 | 78.17 | 0.74ns | 0.008 | 10.40 | 3.08ns | 0.026 | 1.27 | 0.58ns | 0.004 | 482.82 | 0.64ns | 0.006 | 6.55 | 0.03ns | 0.000 | 941.84 | 0.11ns | 0.001 |
| GW2*qSW5 | 1 | 300.02 | 2.00ns | 0.020 | 50.81 | 0.48ns | 0.005 | 2.72 | 0.81ns | 0.007 | 0.17 | 0.08ns | 0.001 | 756.13 | 1.01ns | 0.009 | 108.65 | 0.54ns | 0.004 | 22574.11 | 2.72ns | 0.024 |
| GS3*qSW5 | 1 | 56.82 | 0.38ns | 0.004 | 23.24 | 0.22ns | 0.002 | 25.33 | 7.51** | 0.063 | 3.64 | 1.66ns | 0.013 | 3093.87 | 4.13* | 0.037 | 253.21 | 1.25ns | 0.009 | 4863.55 | 0.59ns | 0.005 |
| GW2*GS3*qSW5 | 1 | 4.85 | 0.03ns | 0.000 | 364.03 | 3.45ns | 0.036 | 0.75 | 0.22ns | 0.002 | 0.07 | 0.03ns | 0.000 | 1746.37 | 2.33ns | 0.021 | 331.32 | 1.64ns | 0.011 | 2586.12 | 0.31ns | 0.003 |
| Model | 7 | 264.27 | 1.76ns | 0.123 | 128.01 | 1.21ns | 0.088 | 15.32 | 4.54** | 0.265 | 13.22 | 6.01** | 0.323 | 2416.32 | 3.22** | 0.204 | 1642.50 | 8.12** | 0.392 | 29736.68 | 3.58** | 0.222 |
| Error | 88 | 150.31 | 105.53 | 3.38 | 2.20 | 749.38 | 202.38 | 8304.46 | ||||||||||||||
| Gene combination | Genotype | n | HDz (DAS) | CL (cm) | PL (cm) | PN | NS | RRG (%) | Yield (g) |
|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||
| Single gene | gw2 | 46 | 100±10.6nsy | 79±11.6ns | 23±2.0ns | 9±1.5** | 110±34.9ns | 53.1±17.94** | 478±96.7** |
| GW2 | 50 | 105±14.2 | 81±9.1 | 23±2.1 | 11±1.6 | 110±26.0 | 72.2±14.36 | 547±96.2 | |
|
|
|||||||||
| gs3 | 42 | 99±11.8* | 82±10.5ns | 24±2.0** | 10±1.8ns | 107±32.3ns | 53.5±18.24** | 479±79.8** | |
| GS3 | 54 | 106±12.9 | 79±10.2 | 22±1.9 | 10±1.9 | 112±29.0 | 70.4±15.64 | 541±109.7 | |
|
|
|||||||||
| qsw5 | 70 | 102±12.2ns | 79±10.8ns | 23±2.0ns | 10±2.0* | 103±30.5** | 61.9±18.43ns | 502±105.6ns | |
| qSW5 | 26 | 104±14.2 | 83±8.8 | 23±2.2 | 9±1.6 | 129±20.2 | 66.1±19.58 | 544±86.4 | |
|
|
|||||||||
| Three gene | gw2gs3qsw5 | 14 | 100±9.2bx | 82±12.6ab | 23±1.9ab | 9±1.2cd | 87±27.6c | 47.9±19.25d | 430±74.2d |
| gw2gs3qSW5 | 9 | 96±6.1b | 78±7.6ab | 25±1.9a | 8±1.1d | 144±21.7a | 44.7±13.97d | 497±68.5bcd | |
| gw2GS3qsw5 | 20 | 102±12.9ab | 77±12.4b | 23±1.9ab | 9±1.8cd | 109±34.8bc | 57.8±16.97cd | 483±96.3cd | |
| gw2GS3qSW5 | 3 | 103±11.3ab | 85±11.5ab | 22±1.5bc | 9±0.6cd | 117±14.3abc | 71.4±9.57abc | 610±153.1a | |
| GW2gs3qsw5 | 13 | 98±12.3b | 80±9.2ab | 24±2.1ab | 11±1.9ab | 97±26.4bc | 58.4±18.34cd | 501±81.3bcd | |
| GW2gs3qSW5 | 6 | 104±21.0ab | 89±9.7a | 25±1.2a | 10±1.0bc | 119±15.2ab | 69.4±7.18bc | 519±62.4abcd | |
| GW2GS3qsw5 | 23 | 106±12.7ab | 80±9.0ab | 22±1.9bc | 11±1.4a | 109±28.3bc | 76.0±7.64ab | 564±111.2abc | |
| GW2GS3qSW5 | 8 | 115±10.8a | 82±6.8ab | 21±0.5c | 11±1.1ab | 124±16.0ab | 85.8±5.38a | 592±63.1ab | |
zHD: heading date, DAS: days after seeding, CL: culm length, PL: panicle length, PN: number of panicles per hill, NS: number of spikelets per panicle, RRG: ratio of ripened grain
yns, *, and ** mean no significant, significant at p<0.05, and p<0.01 by t-test, respectively
xMeans with the same letters in a column are not significantly difference at p<0.05 (ANOVA followed by DMRT)
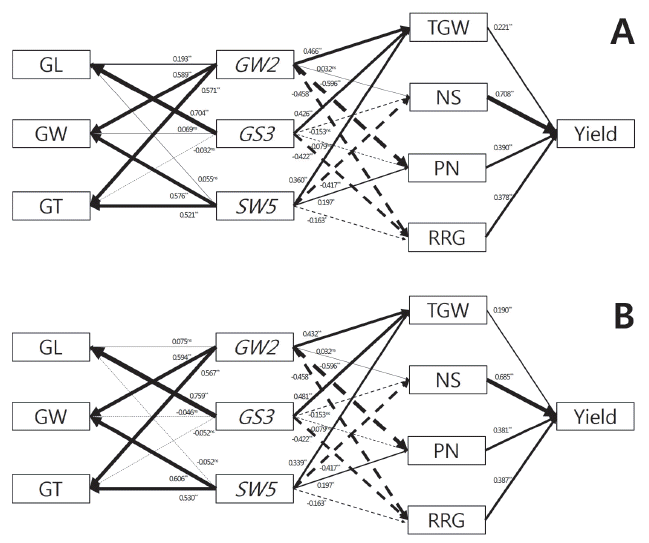

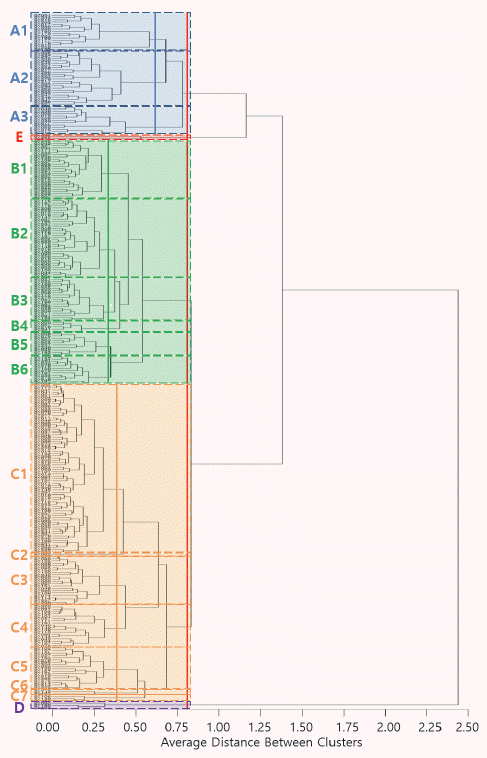
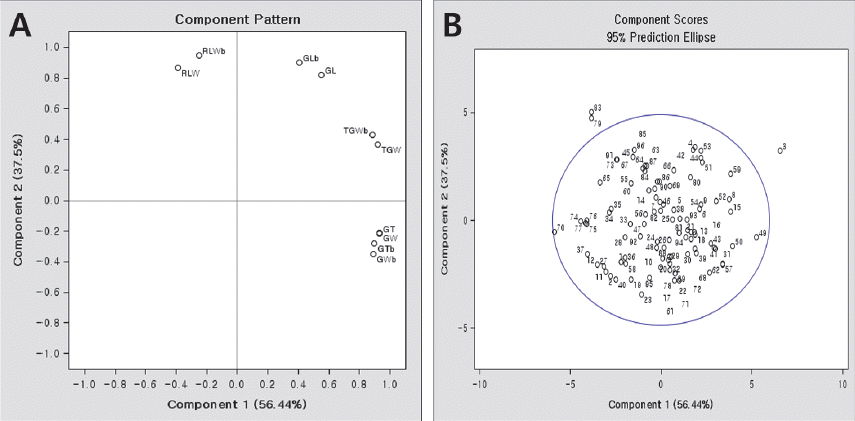
The information of markers used for the identification of grain-related genes, GW2, GS3, and qSW5.
| Gene | Chromosome | Marker | Sequence (5'-3') | Product size (bp) | Reference |
|---|---|---|---|---|---|
|
|
|||||
| GW2 | 2 | GW2-HpaI | F: CCAATAAAGATGTCCATTCTGTTA | 23+151/174 | Yan et al. 2009 |
| R: GCTCTTCCTGTAACACATATTATG | (dCAPS/Hpa I) | Yan et al. 2011 | |||
| GS3 | 3 | GS3-PstI | F: TATTTATTGGCTTGATTTCCTGTG | 294+218/512 | Yan et al. 2009 |
| R: GCTGGTTTTTTACTTTCATTTGCC | (CAPS/Pst I) | Yan et al. 2011 | |||
| qSW5 | 5 | N1212del | F: CGTCTTGCAACCAACGCCGATGTTATAC | 759/1021/1971 | Shomura et al. 2008 |
| R: GAGCGTGTGTAGGGAAGGAGCTGCATGA | (Indel) | Yan et al. 2011 | |||
Anther culture ability of the crosses and doubled haploid lines used in this study.
| Cross combination | Anther cultured (no.) | Cali induced (no. and %) | Plant regeneration (no. and %) |
Green plants (no. and %) |
Albino (no. and %) |
Doubled haploid (DH) lines (no.) |
Selected DH lines (no.) |
|---|---|---|---|---|---|---|---|
|
|
|||||||
| Deuraechan/Jizi1560 | 10,700 | 6,280(58.7) | 1241(19.8) | 146(11.8) | 1095(88.2) | 99 | 88 |
| Deuraechan/Jizi1581 | 11,600 | 2,780(24.0) | 577(20.8) | 110(18.8) | 467(80.9) | 105 | 70 |
| Boramchan/Jizi1560 | 11,800 | 4,320(36.6) | 817(18.9) | 112(13.7) | 705(86.3) | 72 | 62 |
| Boramchan/Jizi1581 | 6,300 | 4,380(69.5) | 1184(27.0) | 228(19.3) | 956(80.7) | 84 | 70 |
| Total | 360 | 290 | |||||
zPercent plant regeneration is the number of plants regenerated over number of cali plated
yPercent green is the number of green plants regenerated over total plants regenerated
xPercent albino is the number of albino plants regenerated over total plants regenerated
wDouble haploid lines are harvested plants having fertile grain
vSelected DH lines are used in this study
Relationship between the grain-related traits of the doubled haploid lines (n=303).
| Trait | GW | GT | RLW | TGW |
|---|---|---|---|---|
|
|
||||
| GL |
0.330** |
0.285** | 0.638** | 0.789** |
| GW | 0.817** | -0.510** | 0.658** | |
| GT | -0.399** | 0.655** | ||
| RLW | 0.174** | |||
zGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
y** means significant at p<0.01
Grain-related and agronomic traits of the groups divided by cluster analysis in 2013.
| Group | n | Rough rice | HD (DAS) | CL (cm) | PL (cm) | PN | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||||
| GL |
GW (mm) | GT (mm) | RLW | TGW (g) | |||||||
|
|
|||||||||||
| A | A1 | 15 | 10.42b |
4.08c | 2.54cd | 2.56c | 46.0b | 96bc | 84abc | 23bcde | 9abcd |
| A2 | 25 | 9.76c | 3.62efg | 2.37fgh | 2.70b | 37.9def | 97bc | 80bc | 24abc | 8bcd | |
| A3 | 13 | 9.69c | 4.46b | 2.68b | 2.18f | 43.3bc | 104bc | 80bc | 23cde | 8cd | |
| Mean | 53 | 9.93C |
3.96B | 2.49B | 2.53B | 41.5B | 98B | 81B | 24AB | 8B | |
| C.V.(%) | 53 | 3.9 | 9.8 | 6.4 | 9.5 | 11.5 | 11.2 | 13.8 | 10.1 | 24.7 | |
| B | B1 | 24 | 8.70f | 3.78ef | 2.40efg | 2.31de | 35.0ef | 96bc | 83abc | 23cde | 9abcd |
| B2 | 36 | 8.79ef | 4.10c | 2.53cd | 2.15f | 37.7def | 99bc | 79bc | 22cde | 8abcd | |
| B3 | 19 | 8.44g | 4.21c | 2.60bc | 2.01g | 37.1def | 101bc | 77bc | 23cde | 8abcd | |
| B4 | 5 | 8.97e | 4.52b | 2.61bc | 1.98gh | 38.3de | 105bc | 72c | 22cde | 8bcd | |
| B5 | 10 | 9.32d | 4.15c | 2.50cde | 2.25ef | 40.1cd | 109b | 89ab | 26a | 7d | |
| B6 | 13 | 9.23d | 3.81de | 2.37fgh | 2.43d | 35.9ef | 95bc | 76bc | 23cde | 9abcd | |
| Mean | 107 | 8.82D | 4.04B | 2.49B | 2.20C | 37.0C | 99B | 80B | 23B | 8B | |
| C.V.(%) | 107 | 3.6 | 5.9 | 4.8 | 7.3 | 9.3 | 13.1 | 15.3 | 10.5 | 21.2 | |
| C | C1 | 73 | 8.10h | 4.01cd | 2.53cd | 2.03g | 34.2f | 97bc | 75bc | 22cde | 9abcd |
| C2 | 1 | 7.59i | 4.05c | 2.42def | 1.88h | 30.2g | 106b | 80bc | 20e | 9abcd | |
| C3 | 22 | 7.77i | 3.63efg | 2.34fgh | 2.15f | 29.3g | 97bc | 75bc | 22cde | 9abcd | |
| C4 | 18 | 7.19j | 3.59fg | 2.34fgh | 2.01g | 27.0g | 100bc | 76bc | 21de | 10abc | |
| C5 | 19 | 8.22gh | 3.44gh | 2.29ghi | 2.40d | 29.8g | 105bc | 83abc | 23bcde | 9abcd | |
| C6 | 2 | 7.68i | 3.16ij | 2.09j | 2.43d | 23.5h | 98cb | 72c | 21e | 10abc | |
| C7 | 3 | 8.80ef | 3.30hi | 2.20i | 2.67bc | 29.9g | 109b | 79bc | 22cde | 9abcd | |
| Mean | 138 | 7.95E | 3.79B | 2.43B | 2.11C | 31.6D | 99B | 76B | 22B | 9AB | |
| C.V.(%) | 138 | 5.0 | 7.9 | 5.8 | 8.5 | 11.7 | 10.1 | 11.8 | 8.7 | 17.8 | |
| D | D | 3 | 11.56aA | 5.26aA | 2.97aA | 2.20efC | 55.3aA | 89cB | 83abcAB | 24abcdAB | 8cdB |
| C.V.(%) | 3 | 1.5 | 3.6 | 1.7 | 2.7 | 2.9 | 0.0 | 3.4 | 8.5 | 13.2 | |
| E | E | 2 | 10.60bB | 3.07jC | 2.27hiC | 3.45aA | 35.0eCD | 125aA | 95aA | 26abA | 11aA |
| C.V.(%) | 2 | 4.8 | 1.6 | 2.6 | 3.2 | 1.2 | 0.0 | 3.3 | 0.8 | 41.7 | |
| Total | Mean | 303 | 8.66 | 3.91 | 2.47 | 2.23 | 35.5 | 99 | 78 | 23 | 9 |
| C.V.(%) | 303 | 10.2 | 9.0 | 6.1 | 11.7 | 16.0 | 12.1 | 13.8 | 10.2 | 20.7 | |
zGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight, HD: heading date, DAS: days after seeding, CL: culm length, PL: panicle length, PN: number of panicles per hill
yMeans with the same letters in a column are not significantly different at p<0.05 (ANOVA followed by DMRT)
xCapital letters indicate statistic difference among main groups defined by culstering analysis
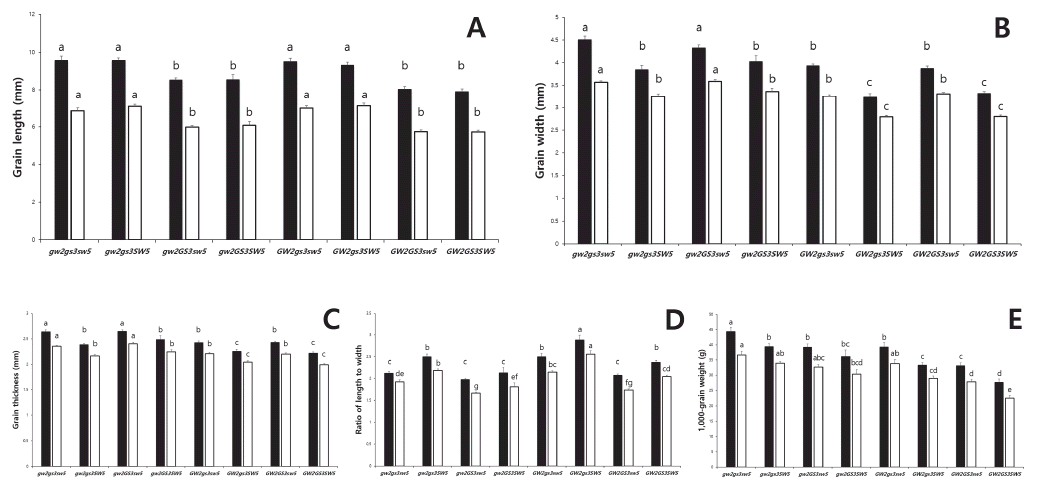
Grain-related traits of the breeding materials.
| Group | n | Rough rice | Brown rice | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
||||||||||||
| GL |
GW (mm) | GT (mm) | RLW | TGW (g) | GL (mm) | GW (mm) | GT (mm) | RLW | TGW (g) | |||
|
|
||||||||||||
| A | A1 | 6 | 10.43b |
4.19cd | 2.57bc | 2.49bc | 47.1b | 7.64b | 3.42bcde | 2.31abcd | 2.24bc | 39.8b |
| A2 | 10 | 9.49c | 3.62ef | 2.37def | 2.63b | 37.7de | 7.04c | 3.09fgh | 2.14cdef | 2.29b | 32.9de | |
| A3 | 8 | 9.53c | 4.53bc | 2.70b | 2.12efg | 45.2bc | 6.81cd | 3.60ab | 2.41a | 1.90def | 37.5bc | |
| Mean | 24 | 9.74B |
4.07B | 2.53B | 2.43B | 42.6B | 7.12B | 3.34A | 2.27AB | 2.15BC | 36.1B | |
| C.V.(%) | 24 | 4.7 | 11.6 | 7.0 | 10.9 | 11.6 | 5.5 | 8.6 | 6.5 | 10.4 | 9.6 | |
| B | B1 | 6 | 8.88d | 3.87de | 2.37ef | 2.30cde | 35.2efg | 6.39de | 3.25cdef | 2.18bcdef | 1.97de | 30.6def |
| B2 | 13 | 8.87d | 4.18cd | 2.56bcd | 2.13defg | 39.7de | 6.32ef | 3.49abcd | 2.31abc | 1.81efg | 33.1de | |
| B3 | 7 | 8.31e | 4.23bcd | 2.54bcde | 1.97fg | 37.1de | 5.96fgh | 3.57ab | 2.35ab | 1.67gh | 31.5de | |
| B4 | 4 | 8.79d | 4.58b | 2.70b | 1.93g | 41.5cd | 6.15efg | 3.72a | 2.41a | 1.66gh | 34.1cd | |
| B5 | 4 | 9.38c | 4.24bcd | 2.51bcde | 2.22cdef | 40.9d | 6.81cd | 3.51abc | 2.22bcde | 1.95de | 34.0cd | |
| B6 | 4 | 9.30c | 3.94de | 2.40cdef | 2.63b | 37.5de | 6.77cd | 3.23def | 2.17bcdef | 2.10bcd | 31.5de | |
| Mean | 38 | 8.86C | 4.16B | 2.52B | 2.17B | 38.6BC | 6.34C | 3.47A | 2.28AB | 1.84CD | 32.4BC | |
| C.V.(%) | 38 | 4.8 | 6.3 | 5.6 | 12.5 | 8.5 | 6.0 | 5.5 | 5.9 | 9.8 | 7.0 | |
| C | C1 | 9 | 8.02ef | 4.17cd | 2.58bc | 1.93g | 35.5ef | 5.64hij | 3.49abcd | 2.35ab | 1.62gh | 29.5ef |
| C2 | 1 | 7.49g | 3.94de | 2.36ef | 1.90g | 29.9h | 5.24jk | 3.34bcdef | 2.13cdef | 1.57h | 24.9ghi | |
| C3 | 5 | 7.58g | 3.64ef | 2.36ef | 2.09efg | 30.1h | 5.46ijk | 3.20efg | 2.12def | 1.71fgh | 25.0ghi | |
| C4 | 5 | 7.25g | 3.66ef | 2.34ef | 1.99fg | 28.0hi | 5.18k | 3.08fgh | 2.13cdef | 1.69gh | 22.6hi | |
| C5 | 6 | 8.06ef | 3.35f | 2.26fg | 2.41bcd | 28.9hi | 5.80ghi | 2.83hi | 2.03ef | 2.05cd | 24.0hi | |
| C6 | 2 | 7.66fg | 3.50f | 2.24gf | 2.22cdef | 24.9i | 5.49ijk | 2.89hi | 1.99f | 1.93de | 21.8i | |
| C7 | 3 | 8.37e | 3.48f | 2.29fg | 2.42bc | 31.1gh | 6.20efg | 2.96gh | 2.07ef | 2.11bcd | 25.9gh | |
| Mean | 31 | 7.83D | 3.73B | 2.39BC | 2.12B | 30.8D | 5.60D | 3.16A | 2.16CB | 1.79D | 25.6D | |
| C.V.(%) | 31 | 5.3 | 9.7 | 6.7 | 10.6 | 13.1 | 6.2 | 9.1 | 7.6 | 12.1 | 13.6 | |
| D | D | 1 | 11.52aA | 5.41aA | 2.97aA | 2.13defgB | 56.6aA | 8.17aA | 3.57abA | 2.45aA | 2.29bB | 48.1aA |
| E | E | 2 | 9.65cB | 3.00gC | 2.14gC | 3.22aA | 32.4fghCD | 7.52bB | 2.69iB | 1.99fC | 2.81aA | 27.9fgCD |
| Total | Mean | 96 | 8.79 | 4.00 | 2.47 | 2.24 | 37.1 | 6.34 | 3.32 | 2.23 | 1.93 | 31.2 |
| C.V.(%) | 96 | 10.2 | 11.2 | 7.3 | 13.9 | 17.2 | 11.5 | 8.9 | 7.2 | 14.5 | 17.3 | |
zGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
yMeans with the same letters in a column are not significantly different at p<0.05 (ANOVA followed by DMRT)
xCapital letters indicate statistic difference among main groups defined by culstering analysis
Eigenvectors for the principal component analysis of the traits and correlation coefficient between the traits and principal components.
| Trait | Eigenvector | Correlation coefficient | ||||
|---|---|---|---|---|---|---|
|
|
||||||
| Prin1 |
Prin2 | Prin3 | Prin1 | Prin2 | Prin3 | |
|
|
||||||
| GL |
0.232 | 0.425 | -0.171 | 0.551** |
0.822** | -0.089ns |
| GW | 0.392 | -0.109 | -0.411 | 0.931** | -0.212* | -0.213* |
| GT | 0.393 | -0.112 | 0.422 | 0.933** | -0.218* | 0.218* |
| RLW | -0.165 | 0.447 | 0.280 | -0.391** | 0.865** | 0.145ns |
| TGW | 0.386 | 0.189 | -0.028 | 0.918** | 0.365** | -0.014ns |
| GLb |
0.170 | 0.466 | -0.180 | 0.405** | 0.902** | -0.093ns |
| GWb | 0.375 | -0.179 | -0.320 | 0.890** | -0.347** | -0.166ns |
| GTb | 0.376 | -0.143 | 0.627 | 0.894** | -0.277** | 0.325** |
| RLWb | -0.106 | 0.491 | 0.122 | -0.253* | 0.950** | 0.063ns |
| TGWb | 0.373 | 0.223 | 0.052 | 0.887** | 0.431** | 0.027ns |
| Eigen value | 5.64 | 3.75 | 0.27 | |||
| Contribution (%) | 56.4 | 37.5 | 2.7 | |||
| Cumulative contribution (%) | 56.4 | 93.9 | 96.6 | |||
zGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
yPrin1, Prin2, and Prin3 indicate the first, second, and third component, respectively
xns, *, and ** mean no significant, significant at p<0.05, and p<0.01, respectively
wb means brown rice
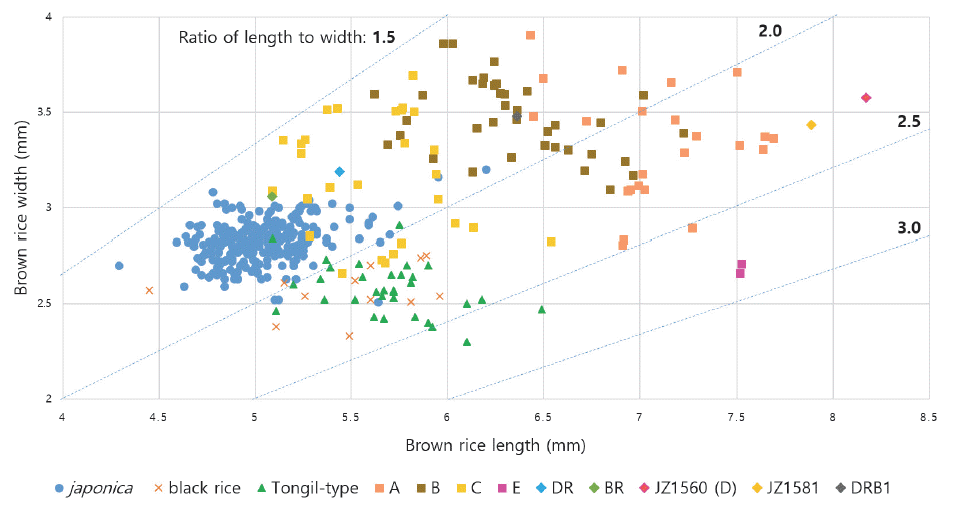
Comparison of grain-related traits of brown rice between the breeding materials and Korean rice cultivars.
| Group | n | GL |
GW (mm) | GT (mm) | RLW | TGW (g) |
|---|---|---|---|---|---|---|
|
|
||||||
| Deuraechan | 1 | 5.44 | 3.19 | 2.12 | 1.71 | 24.7 |
| Boramchan | 1 | 5.09 | 3.06 | 2.13 | 1.67 | 22.9 |
| Jizi1560 | 1 | 8.17 | 3.57 | 2.45 | 2.29 | 48.1 |
| Jizi1581 | 1 | 7.89 | 3.43 | 2.29 | 2.30 | 38.0 |
| Daeripbyeo 1 | 1 | 6.36 | 3.48 | 2.25 | 1.83 | 34.6 |
| DH lines | 91 | 6.34±0.68a |
3.32±0.30a | 2.23±0.16a | 1.93±0.28c | 31.1±5.1a |
| (5.09-7.69) | (2.66-3.91) | (1.83-2.63) | (1.53-2.83) | (18.6-41.5) | ||
| japonica | 264 | 5.03±0.23d |
2.82±0.11b | 1.94±0.13b | 1.79±0.09d | 22.0±2.0b |
| (4.29-6.20) | (2.51-3.20) | (1.23-2.30) | (1.55-2.02) | (14.1-34.8) | ||
| black rice | 13 | 5.34±0.63c | 2.57±0.12c | 1.78±0.11c | 2.08±0.25b | 19.5±2.9c |
| (3.76-5.96) | (2.33-2.75) | (1.57-1.97) | (1.46-2.36) | (14.8-23.5) | ||
| Tongil-type | 32 | 5.69±0.30b | 2.57±0.13c | 1.83±0.13c | 2.22±0.19a | 22.5±2.1b |
| (5.09-6.49) | (2.30-2.91) | (1.60-2.14) | (1.79-2.65) | (18.9-28.2) | ||
zGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
yMeans with the same letters in a column are not significantly different at p<0.05 (ANOVA followed by DMRT)
xThe data of japonica, black rice, and Tongil-type cultivars were used the value on registration of national variety and in the website of 'Nongsaro' (http://www.nongsaro.go.kr/)
Yield-related traits of the breeding materials.
| Group | n | HD |
CL (cm) | PL (cm) | PN | NS | RRG (%) | Yield (g) | |
|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||
| A | A1 | 6 | 95bc |
80ab | 24abc | 11abc | 68de | 66.6abcd | 468cdef |
| A2 | 10 | 94bc | 80ab | 26a | 9bc | 136ab | 52.7de | 494bcdef | |
| A3 | 8 | 102bc | 84ab | 23abc | 9abc | 88cde | 39.9e | 405ef | |
| Mean | 24 | 97B |
82B | 24A | 9A | 103AB | 51.9A | 458AB | |
| C.V.(%) | 24 | 10.5 | 13.9 | 8.2 | 19.6 | 38.2 | 36.2 | 16.7 | |
| B | B1 | 6 | 97bc | 79b | 24abc | 11abc | 95bcde | 55.7cde | 483bcdef |
| B2 | 13 | 102bc | 80ab | 23abc | 10abc | 94bcde | 68.2abcd | 511abcdef | |
| B3 | 7 | 106bc | 81ab | 23abc | 8c | 125abc | 59.8bcde | 527abcde | |
| B4 | 4 | 107bc | 73b | 22bc | 10abc | 93bcde | 53.4de | 453def | |
| B5 | 4 | 109abc | 90ab | 24abc | 8c | 112abc | 54.1de | 446def | |
| B6 | 4 | 98bc | 82ab | 25ab | 11abc | 105abcd | 56.5cde | 544abcde | |
| Mean | 38 | 103B | 81B | 23A | 10A | 103AB | 60.4A | 500AB | |
| C.V.(%) | 38 | 13.2 | 14.4 | 8.3 | 19.5 | 24.2 | 26.2 | 17.9 | |
| C | C1 | 9 | 99bc | 75b | 23abc | 10abc | 118abc | 64.7abcde | 523abcdef |
| C2 | 1 | 109abc | 83ab | 21c | 12a | 123abc | 84.6ab | 632abc | |
| C3 | 5 | 109abc | 76b | 22bc | 12ab | 118abc | 77.5abcd | 554abcde | |
| C4 | 5 | 104bc | 76b | 21c | 12ab | 135ab | 81.5abc | 641ab | |
| C5 | 6 | 115ab | 83ab | 21c | 10abc | 125abc | 86.9a | 596abcd | |
| C6 | 2 | 104bc | 73b | 22bc | 11abc | 150a | 69.1abcd | 666a | |
| C7 | 3 | 110abc | 81ab | 23abc | 10abc | 122abc | 78.8abcd | 556abcde | |
| Mean | 31 | 106B | 78B | 22A | 11A | 125A | 76.1A | 577A | |
| C.V.(%) | 31 | 10.3 | 8.2 | 8.1 | 18.0 | 18.5 | 15.4 | 17.8 | |
| D | D | 1 | 92cB | 91abAB | 24abcA | 10abcA | 55eB | 7.1fB | 362fB |
| E | E | 2 | 128aA | 98aA | 24abcA | 10abcA | 123abcA | 73.8abcdA | 542abcdeA |
| Total | Mean | 96 | 103 | 80 | 23 | 10 | 110 | 63.0 | 513 |
| C.V.(%) | 96 | 12.4 | 12.9 | 8.8 | 18.4 | 27.7 | 29.7 | 19.9 | |
zHD: heading date, DAS: days after seeding, CL: culm length, PL: panicle length, PN: number of panicles per hill, NS: number of spikelets per panicle, RRG: ratio of ripened grain
yMeans with the same letters in a column are not significantly different at p<0.05 (ANOVA followed by DMRT)
xCapital letters indicate statistic difference among main groups defined by culstering analysis
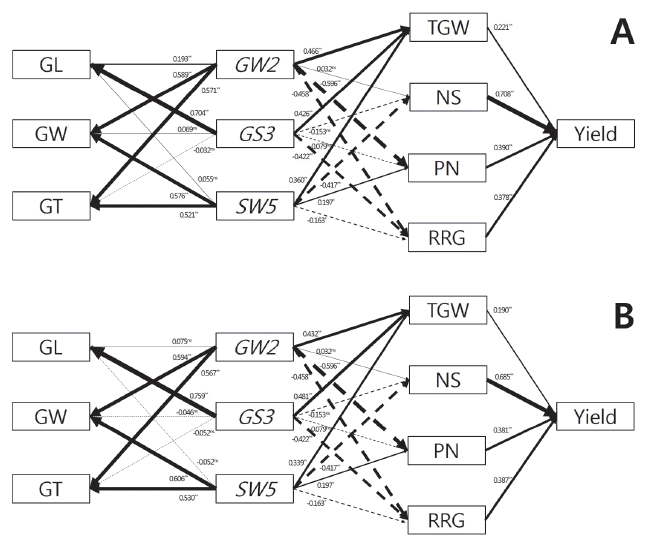
Correlation coefficient among grain and yield-related traits in the breeding materials.
| Trait |
GL |
GW | GT | RLW | TGW | HD | CL | PL | PN | NS | RRG | Yield |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (x1, 6) | (x2, 7) | (x3, 8) | (x4, 9) | (x5, 10) | (x11) | (x12) | (x13) | (x14) | (x15) | (x16) | (x17) | |
|
|
||||||||||||
| x1, 6 | 0.364** | 0.322** | 0.478** | 0.801** | -0.255* | 0.237* | 0.475** | -0.168ns | -0.427** | -0.508** | -0.476** | |
| x2, 7 | 0.066ns |
0.887** | -0.576** | 0.768** | -0.310** | -0.127ns | 0.043ns | -0.230* | -0.463** | -0.561** | -0.491** | |
| x3, 8 | 0.088ns | 0.850** | -0.534** | 0.770** | -0.190ns | -0.068ns | 0.038ns | -0.249* | -0.431** | -0.417** | -0.427** | |
| x4, 8 | 0.759** | -0.591** | -0.467** | -0.043ns | 0.075ns | 0.255* | 0.396** | 0.037ns | 0.048ns | 0.080ns | 0.012ns | |
| x5, 10 | 0.741** | 0.639** | 0.680** | 0.177ns | -0.270** | 0.103ns | 0.292** | -0.213* | -0.576** | -0.498** | -0.554** | |
| x11 | -0.220* | -0.208* | -0.166ns | -0.024ns | -0.276** | 0.525** | -0.257* | -0.007ns | 0.290** | 0.440** | 0.434** | |
| x12 | 0.289** | -0.133ns | -0.090ns | 0.324** | 0.106 | 0.525** | 0.141ns | -0.192ns | 0.108ns | 0.115ns | 0.209* | |
| x13 | 0.477** | 0.057ns | 0.033ns | 0.355** | 0.366** | -0.257* | 0.141ns | -0.425** | 0.132ns | -0.324** | -0.274** | |
| x14 | -0.150ns | -0.269** | -0.285** | 0.033ns | -0.243* | -0.007ns | -0.192ns | -0.425** | -0.451** | 0.462** | 0.240* | |
| x15 | -0.345** | -0.381** | -0.359** | -0.022ns | -0.506** | 0.290** | 0.108ns | 0.132ns | -0.451** | 0.038ns | 0.507** | |
| x16 | -0.447** | -0.398** | -0.375** | -0.100ns | -0.522** | 0.440** | 0.115ns | -0.324** | 0.462** | 0.038ns | 0.575** | |
| x17 | -0.420** | -0.410** | -0.367** | -0.081ns | -0.529** | 0.434** | 0.209* | -0.274** | 0.240* | 0.507** | 0.575** | |
zUpper and lower diagonal indicate the value of rough and brown rice, respectively
yGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight, HD: heading date, CL: culm length, PL: panicle length, PN: number of panicles per hill, NS: number of spikelets per panicle, RRG: ratio of ripened grain
xns, *, and ** mean no significant, significant at p<0.05, and p<0.01, respectively
Number of allele types and frequency of GW2, GS3, and qSW5 in each group of the breeding materials.
| Group | n | GW2 | GS3 | qSW5 | Remark | ||||
|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||
| gw2 | GW2 | gs3 | GS3 | qsw5 | qSW5 | ||||
|
|
|||||||||
| A1 | 6 | 3 | 3 | 6 | 0 | 5 | 1 | Jizi1581 (gw2gs3qSW5) | |
| A2 | 10 | 6 | 4 | 10 | 0 | 1 | 9 | ||
| A | A3 | 8 | 7 | 1 | 6 | 2 | 8 | 0 | |
| Total | 24 | 16 | 8 | 22 | 2 | 14 | 10 | ||
| (66.7) |
(33.3) | (91.7) | (8.3) | (58.3) | (41.7) | ||||
| B1 | 6 | 0 | 6 | 3 | 3 | 6 | 0 | Daeripbyeo 1 (gw2GS3qsw5) | |
| B2 | 13 | 9 | 4 | 3 | 10 | 11 | 2 | ||
| B3 | 7 | 6 | 1 | 2 | 5 | 7 | 0 | ||
| B | B4 | 4 | 4 | 0 | 1 | 3 | 4 | 0 | |
| B5 | 4 | 3 | 1 | 3 | 1 | 3 | 1 | ||
| B6 | 4 | 1 | 3 | 3 | 1 | 3 | 1 | ||
| Total | 38 | 23 | 15 | 15 | 23 | 34 | 4 | ||
| (60.5) | (39.5) | (39.5) | (60.5) | (89.5) | (10.5) | ||||
| C1 | 9 | 5 | 4 | 0 | 9 | 8 | 1 | ||
| C2 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | Deuraechan (GW2GS3qsw5) | |
| C3 | 5 | 0 | 5 | 0 | 5 | 5 | 0 | ||
| C4 | 5 | 0 | 5 | 0 | 5 | 4 | 1 | Boramchan (GW2GS3qsw5) | |
| C | C5 | 6 | 0 | 6 | 1 | 5 | 1 | 5 | |
| C6 | 2 | 1 | 1 | 0 | 2 | 1 | 1 | ||
| C7 | 3 | 0 | 3 | 1 | 2 | 1 | 2 | ||
| Total | 31 | 6 | 25 | 2 | 29 | 21 | 10 | ||
| (19.4) | (80.6) | (6.5) | (93.5) | (67.7) | (32.3) | ||||
| D | D | 1 | 1 | 0 | 1 | 0 | 1 | 0 | Jizi1560 (gw2gs3qsw5) |
| (100) | (0) | (100) | (0) | (100) | (0) | ||||
| E | E | 2 | 0 | 2 | 2 | 0 | 0 | 2 | DGS79, 83 (GW2gs3qSW5) |
| (0) | (100) | (100) | (0) | (0) | (100) | ||||
| Total | 96 | 46 | 50 | 42 | 54 | 70 | 26 | ||
| (47.9) | (52.1) | (43.8) | (56.3) | (72.9) | (27.1) | ||||
zThe value in the parenthesis is the frequency of the allele type of GW2, GS3, and qSW5
The influence of GW2, GS3, and qSW5 on the phenotypic variation of grain-related traits.
| Source of variance | df | Rough rice | Brown rice | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||||||||||||||||||||||||
| GL |
GW | GT | RLW | TGW | GL | GW | GT | RLW | TGW | ||||||||||||||||||||||
|
|
|||||||||||||||||||||||||||||||
| MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | MS | F | R 2 | ||
|
|
|||||||||||||||||||||||||||||||
| GW2 | 1 | 2.13 | 5.72* |
0.035 | 5.61 | 90.34** | 0.340 | 0.71 | 53.57** | 0.276 | 1.26 | 39.77** | 0.156 | 668.91 | 32.63** | 0.202 | 0.19 | 0.92ns | 0.005 | 2.72 | 123.16** | 0.372 | 0.54 | 53.22** | 0.268 | 0.83 | 47.10** | 0.131 | 431.83 | 30.77** | 0.182 |
| GS3 | 1 | 25.04 | 67.36** | 0.411 | 0.00 | 0.01ns | 0.000 | 0.01 | 0.50ns | 0.003 | 2.15 | 67.75** | 0.266 | 417.16 | 20.35** | 0.126 | 20.71 | 98.71** | 0.513 | 0.04 | 1.70ns | 0.005 | 0.01 | 0.55ns | 0.003 | 2.43 | 137.72** | 0.383 | 412.70 | 29.41** | 0.174 |
| qSW5 | 1 | 0.10 | 0.26ns | 0.002 | 5.02 | 80.83** | 0.304 | 0.64 | 48.78** | 0.251 | 1.54 | 48.57** | 0.191 | 387.05 | 18.88** | 0.117 | 0.21 | 0.99ns | 0.005 | 2.39 | 108.30** | 0.327 | 0.54 | 52.58** | 0.265 | 1.29 | 73.23** | 0.204 | 235.22 | 16.76** | 0.099 |
| GW2*GS3 | 1 | 0.73 | 1.97ns | 0.012 | 0.00 | 0.02ns | 0.000 | 0.02 | 1.49ns | 0.008 | 0.18 | 5.65* | 0.022 | 11.85 | 0.58ns | 0.004 | 0.62 | 2.97ns | 0.015 | 0.01 | 0.28ns | 0.001 | 0.04 | 3.76ns | 0.019 | 0.09 | 5.00* | 0.014 | 25.40 | 1.81ns | 0.011 |
| GW2*qSW5 | 1 | 0.12 | 0.33ns | 0.002 | 0.09 | 1.41ns | 0.005 | 0.00 | 0.02ns | 0.000 | 0.03 | 0.89ns | 0.003 | 12.64 | 0.62ns | 0.004 | 0.06 | 0.29ns | 0.002 | 0.19 | 8.70** | 0.026 | 0.00 | 0.02ns | 0.000 | 0.10 | 5.52* | 0.015 | 27.71 | 1.97ns | 0.012 |
| GS3*qSW5 | 1 | 0.01 | 0.02ns | 0.000 | 0.26 | 4.17* | 0.016 | 0.00 | 0.22ns | 0.001 | 0.10 | 3.03ns | 0.012 | 5.44 | 0.27ns | 0.002 | 0.08 | 0.40ns | 0.002 | 0.00 | 0.13ns | 0.000 | 0.00 | 0.01ns | 0.000 | 0.05 | 2.84ns | 0.008 | 0.04 | 0.00ns | 0.000 |
| GW2*GS3*qSW5 | 1 | 0.00 | 0.00 ns | 0.000 | 0.05 | 0.79 ns | 0.003 | 0.02 | 1.46 ns | 0.008 | 0.02 | 0.57 ns | 0.002 | 2.06 | 0.10 ns | 0.001 | 0.00 | 0.00 ns | 0.000 | 0.02 | 0.82 ns | 0.002 | 0.01 | 0.59 ns | 0.003 | 0.00 | 0.01 ns | 0.000 | 0.36 | 0.03 ns | 0.000 |
| Model | 7 | 4.02 | 10.81** | 0.462 | 1.57 | 25.37** | 0.669 | 0.20 | 15.15** | 0.546 | 0.75 | 23.75** | 0.654 | 215.01 | 10.49** | 0.455 | 3.13 | 14.90** | 0.542 | 0.77 | 34.73** | 0.734 | 0.16 | 15.82** | 0.557 | 0.68 | 38.77** | 0.755 | 161.89 | 11.54** | 0.479 |
| Error | 88 | 0.372 | 0.062 | 0.013 | 0.032 | 20.50 | 0.210 | 0.022 | 0.010 | 0.018 | 14.03 | ||||||||||||||||||||
zGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
2)ns, *, and ** mean no significant, significant at p<0.05, and p<0.01 by three-way ANOVA, respectively
Comparison of the value of grain-related traits according to allele types.
| Gene combination | Genotype | n | Rough rice | Brown rice | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
||||||||||||
| GL |
GW (mm) | GT (mm) | RLW | TGW (g) | GL (mm) | GW (mm) | GT (mm) | RLW | TGW (g) | |||
|
|
||||||||||||
| Single gene | gw2 | 46 | 9.05±0.797** |
4.27±0.373** | 2.58±0.159** | 2.14±0.240** | 40.6±5.31** | 6.46±0.656ns | 3.50±0.211** | 2.33±0.139** | 1.86±0.237* | 34.0±4.16** |
| GW2 | 50 | 8.55±0.921 | 3.73±0.341 | 2.38±0.136 | 2.33±0.340 | 33.9±5.61 | 6.23±0.774 | 3.15±0.260 | 2.15±0.127 | 1.99±0.303 | 28.7± 5.15 | |
| gs3 | 42 | 9.51±0.633** | 4.01±0.434ns | 2.46±0.174ns | 2.43±0.339** | 40.2±5.65** | 6.98±0.494** | 3.34±0.281ns | 2.22±0.132ns | 2.14±0.255** | 34.1±4.46** | |
| GS3 | 54 | 8.23±0.621 | 3.97±0.421 | 2.48±0.185 | 2.09±0.180 | 34.8±5.97 | 5.84±0.431 | 3.29±0.308 | 2.25±0.181 | 1.76±0.166 | 29.0±4.99 | |
| qsw5 | 70 | 8.76±0.921ns | 4.14±0.366** | 2.53±0.161** | 2.14±0.250** | 38.3±6.33** | 6.26±0.714ns | 3.43±0.213** | 2.29±0.141** | 1.83±0.218** | 32.1±5.25** | |
| qSW5 | 26 | 8.88±0.829 | 3.56±0.372 | 2.32± 0.125 | 2.51±0.298 | 34.0±5.57 | 6.55±0.729 | 3.02±0.276 | 2.09±0.117 | 2.19±0.269 | 28.9±5.15 | |
| Three genes | gw2gs3qsw5 | 14 | 9.57±0.852a |
4.51±0.296a | 2.64±0.133a | 2.13±0.160c | 44.4±5.45a | 6.84±0.600a | 3.57±0.127a | 2.35±0.070a | 1.92±0.185de | 36.6±4.87a |
| gw2gs3qSW5 | 9 | 9.56±0.433a | 3.85±0.274b | 2.39±0.069b | 2.50±0.192b | 39.4±2.82b | 7.08±0.363a | 3.25±0.164b | 2.16±0.088 | 2.18±0.121b | 34.0±1.55ab | |
| gw2GS3qsw5 | 20 | 8.53±0.507b | 4.33±0.305a | 2.65±0.137a | 1.98±0.095c | 39.2±5.08b | 5.98±0.384b | 3.59±0.191a | 2.40±0.132a | 1.67±0.095g | 32.7±3.76abc | |
| gw2GS3qSW5 | 3 | 8.55±0.475b | 4.03±0.220b | 2.49±0.122b | 2.13±0.214c | 36.2±3.67bc | 6.09±0.361b | 3.36±0.123b | 2.24±0.072b | 1.81±0.150ef | 30.3±2.75bcd | |
| GW2gs3qsw5 | 13 | 9.51±0.606a | 3.93±0.155b | 2.43±0.120b | 2.50±0.297b | 39.3±5.00b | 6.98±0.503a | 3.26±0.104b | 2.20±0.087b | 2.15±0.119bc | 33.9±4.31ab | |
| GW2gs3qSW5 | 6 | 9.32±0.393a | 3.24±0.189c | 2.25±0.102c | 2.89±0.258a | 33.4±2.10c | 7.11±0.391a | 2.79±0.085c | 2.04±0.066c | 2.56±0.203a | 29.0±1.98cd | |
| GW2GS3qsw5 | 23 | 8.03±0.680b | 3.88±0.242b | 2.43±0.106b | 2.08±0.141c | 33.2±4.53c | 5.74±0.492b | 3.31±0.161b | 2.20±0.112b | 1.74±0.126gf | 27.8±3.94d | |
| GW2GS3qSW5 | 8 | 7.90±0.393b | 3.32±0.122c | 2.22±0.069c | 2.38±0.097b | 27.7±3.00d | 5.72±0.279b | 2.80±0.092c | 1.99±0.077c | 2.05±0.082cd | 22.5±2.39e | |
zGL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
yns, *, and ** mean no significant, significant at p<0.05, and p<0.01 by t-test, respectively
xMeans with the same letters in a column are not significantly difference at p<0.05 (ANOVA followed by DMRT)
The influence of GW2, GS3, and qSW5 on the phenotypic variation of yield-related traits.
| Source of variance | df | HD |
CL | PL | PN | NS | RRG | Yield | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
||||||||||||||||||||||
| MS | MS | R 2 | MS | MS | R 2 | MS | MS | R 2 | MS | MS | R 2 | MS | MS | R 2 | MS | MS | R 2 | MS | MS | R 2 | ||
|
|
||||||||||||||||||||||
| GW2 | 1 | 456.17 | 3.03ns |
0.030 | 69.24 | 0.66ns | 0.007 | 1.92 | 0.57ns | 0.005 | 71.89 | 32.69** | 0.251 | 68.69 | 0.09ns | 0.001 | 4710.60 | 23.28** | 0.161 | 25221.91 | 3.04ns | 0.027 |
| GS3 | 1 | 771.18 | 5.13* | 0.051 | 23.78 | 0.23ns | 0.002 | 65.81 | 19.50** | 0.163 | 4.44 | 2.02ns | 0.016 | 154.29 | 0.21ns | 0.002 | 5093.57 | 25.17** | 0.174 | 92956.71 | 11.19** | 0.099 |
| qSW5 | 1 | 149.60 | 1.00ns | 0.010 | 286.80 | 2.72ns | 0.028 | 0.29 | 0.09ns | 0.001 | 11.05 | 5.02* | 0.039 | 10612.05 | 14.16** | 0.128 | 993.58 | 4.91* | 0.034 | 59012.53 | 7.11** | 0.063 |
| GW2*GS3 | 1 | 111.21 | 0.74ns | 0.007 | 78.17 | 0.74ns | 0.008 | 10.40 | 3.08ns | 0.026 | 1.27 | 0.58ns | 0.004 | 482.82 | 0.64ns | 0.006 | 6.55 | 0.03ns | 0.000 | 941.84 | 0.11ns | 0.001 |
| GW2*qSW5 | 1 | 300.02 | 2.00ns | 0.020 | 50.81 | 0.48ns | 0.005 | 2.72 | 0.81ns | 0.007 | 0.17 | 0.08ns | 0.001 | 756.13 | 1.01ns | 0.009 | 108.65 | 0.54ns | 0.004 | 22574.11 | 2.72ns | 0.024 |
| GS3*qSW5 | 1 | 56.82 | 0.38ns | 0.004 | 23.24 | 0.22ns | 0.002 | 25.33 | 7.51** | 0.063 | 3.64 | 1.66ns | 0.013 | 3093.87 | 4.13* | 0.037 | 253.21 | 1.25ns | 0.009 | 4863.55 | 0.59ns | 0.005 |
| GW2*GS3*qSW5 | 1 | 4.85 | 0.03ns | 0.000 | 364.03 | 3.45ns | 0.036 | 0.75 | 0.22ns | 0.002 | 0.07 | 0.03ns | 0.000 | 1746.37 | 2.33ns | 0.021 | 331.32 | 1.64ns | 0.011 | 2586.12 | 0.31ns | 0.003 |
| Model | 7 | 264.27 | 1.76ns | 0.123 | 128.01 | 1.21ns | 0.088 | 15.32 | 4.54** | 0.265 | 13.22 | 6.01** | 0.323 | 2416.32 | 3.22** | 0.204 | 1642.50 | 8.12** | 0.392 | 29736.68 | 3.58** | 0.222 |
| Error | 88 | 150.31 | 105.53 | 3.38 | 2.20 | 749.38 | 202.38 | 8304.46 | ||||||||||||||
zHD: heading date, CL: culm length, PL: panicle length, PN: number of panicles per hill, NS: number of spikelets per panicle, RRG: ratio of ripened grain
yns, *, and ** mean no significant, significant at p<0.05, and p<0.01 by three-way ANOVA, respectively
Comparison of the value of yield-related traits according to allele types.
| Gene combination | Genotype | n | HD |
CL (cm) | PL (cm) | PN | NS | RRG (%) | Yield (g) |
|---|---|---|---|---|---|---|---|---|---|
|
|
|||||||||
| Single gene | gw2 | 46 | 100±10.6ns |
79±11.6ns | 23±2.0ns | 9±1.5** | 110±34.9ns | 53.1±17.94** | 478±96.7** |
| GW2 | 50 | 105±14.2 | 81±9.1 | 23±2.1 | 11±1.6 | 110±26.0 | 72.2±14.36 | 547±96.2 | |
| gs3 | 42 | 99±11.8* | 82±10.5ns | 24±2.0** | 10±1.8ns | 107±32.3ns | 53.5±18.24** | 479±79.8** | |
| GS3 | 54 | 106±12.9 | 79±10.2 | 22±1.9 | 10±1.9 | 112±29.0 | 70.4±15.64 | 541±109.7 | |
| qsw5 | 70 | 102±12.2ns | 79±10.8ns | 23±2.0ns | 10±2.0* | 103±30.5** | 61.9±18.43ns | 502±105.6ns | |
| qSW5 | 26 | 104±14.2 | 83±8.8 | 23±2.2 | 9±1.6 | 129±20.2 | 66.1±19.58 | 544±86.4 | |
| Three gene | gw2gs3qsw5 | 14 | 100±9.2b |
82±12.6ab | 23±1.9ab | 9±1.2cd | 87±27.6c | 47.9±19.25d | 430±74.2d |
| gw2gs3qSW5 | 9 | 96±6.1b | 78±7.6ab | 25±1.9a | 8±1.1d | 144±21.7a | 44.7±13.97d | 497±68.5bcd | |
| gw2GS3qsw5 | 20 | 102±12.9ab | 77±12.4b | 23±1.9ab | 9±1.8cd | 109±34.8bc | 57.8±16.97cd | 483±96.3cd | |
| gw2GS3qSW5 | 3 | 103±11.3ab | 85±11.5ab | 22±1.5bc | 9±0.6cd | 117±14.3abc | 71.4±9.57abc | 610±153.1a | |
| GW2gs3qsw5 | 13 | 98±12.3b | 80±9.2ab | 24±2.1ab | 11±1.9ab | 97±26.4bc | 58.4±18.34cd | 501±81.3bcd | |
| GW2gs3qSW5 | 6 | 104±21.0ab | 89±9.7a | 25±1.2a | 10±1.0bc | 119±15.2ab | 69.4±7.18bc | 519±62.4abcd | |
| GW2GS3qsw5 | 23 | 106±12.7ab | 80±9.0ab | 22±1.9bc | 11±1.4a | 109±28.3bc | 76.0±7.64ab | 564±111.2abc | |
| GW2GS3qSW5 | 8 | 115±10.8a | 82±6.8ab | 21±0.5c | 11±1.1ab | 124±16.0ab | 85.8±5.38a | 592±63.1ab | |
zHD: heading date, DAS: days after seeding, CL: culm length, PL: panicle length, PN: number of panicles per hill, NS: number of spikelets per panicle, RRG: ratio of ripened grain
yns, *, and ** mean no significant, significant at p<0.05, and p<0.01 by t-test, respectively
xMeans with the same letters in a column are not significantly difference at p<0.05 (ANOVA followed by DMRT)
Percent plant regeneration is the number of plants regenerated over number of cali plated
Percent green is the number of green plants regenerated over total plants regenerated
Percent albino is the number of albino plants regenerated over total plants regenerated
Double haploid lines are harvested plants having fertile grain
Selected DH lines are used in this study
GL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
** means significant at p<0.01
GL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight, HD: heading date, DAS: days after seeding, CL: culm length, PL: panicle length, PN: number of panicles per hill
Means with the same letters in a column are not significantly different at p<0.05 (ANOVA followed by DMRT)
Capital letters indicate statistic difference among main groups defined by culstering analysis
GL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
Means with the same letters in a column are not significantly different at p<0.05 (ANOVA followed by DMRT)
Capital letters indicate statistic difference among main groups defined by culstering analysis
GL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
Prin1, Prin2, and Prin3 indicate the first, second, and third component, respectively
ns, *, and ** mean no significant, significant at p<0.05, and p<0.01, respectively
b means brown rice
GL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
Means with the same letters in a column are not significantly different at p<0.05 (ANOVA followed by DMRT)
The data of japonica, black rice, and Tongil-type cultivars were used the value on registration of national variety and in the website of 'Nongsaro' (http://www.nongsaro.go.kr/)
HD: heading date, DAS: days after seeding, CL: culm length, PL: panicle length, PN: number of panicles per hill, NS: number of spikelets per panicle, RRG: ratio of ripened grain
Means with the same letters in a column are not significantly different at p<0.05 (ANOVA followed by DMRT)
Capital letters indicate statistic difference among main groups defined by culstering analysis
Upper and lower diagonal indicate the value of rough and brown rice, respectively
GL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight, HD: heading date, CL: culm length, PL: panicle length, PN: number of panicles per hill, NS: number of spikelets per panicle, RRG: ratio of ripened grain
ns, *, and ** mean no significant, significant at p<0.05, and p<0.01, respectively
The value in the parenthesis is the frequency of the allele type of GW2, GS3, and qSW5
GL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
ns, *, and ** mean no significant, significant at p<0.05, and p<0.01 by three-way ANOVA, respectively
GL: grain length, GW: grain width, GT: grain thickness, RLW: ratio of length to width, TGW: 1000-grain weight
ns, *, and ** mean no significant, significant at p<0.05, and p<0.01 by t-test, respectively
Means with the same letters in a column are not significantly difference at p<0.05 (ANOVA followed by DMRT)
HD: heading date, CL: culm length, PL: panicle length, PN: number of panicles per hill, NS: number of spikelets per panicle, RRG: ratio of ripened grain
ns, *, and ** mean no significant, significant at p<0.05, and p<0.01 by three-way ANOVA, respectively
HD: heading date, DAS: days after seeding, CL: culm length, PL: panicle length, PN: number of panicles per hill, NS: number of spikelets per panicle, RRG: ratio of ripened grain
ns, *, and ** mean no significant, significant at p<0.05, and p<0.01 by t-test, respectively
Means with the same letters in a column are not significantly difference at p<0.05 (ANOVA followed by DMRT)

