적요
Drought stress during the seedling stage has a disastrous effect on the growth of maize. The purpose of this study was to assess the developed expression markers that are related to drought stress in maize. For the selection of expressed genes by drought stress, co-expression analysis was carried out using published microarray data of drought stress in maize (Zea mays L.) seedlings. Six consensus modules were based on 4,770 stress responsive genes differentially expressed in drought stress, and the royal blue module was chosen. Thirty genes were selected according to different expression patterns between susceptible and tolerant types. Drought stress treatments were performed on both Ki3 and Ki11. Ki3 and Ki11 are widely known drought-susceptible and -tolerant types, respectively. At first, the 30 selected genes were compared to Ki3 and Ki11 using qRT-PCR. The gene expression values of eight genes (BU050895, BF728598, CK827168, CO524848, AF457983, CF037152, AJ606944, and BG836522) were significantly tolerant types rather than susceptible types in the roots. After applying the eight above-mentioned genes to nine cultivars, a different pattern was detected between susceptible and tolerant types. The results of the present study will show the possibility of developing novel expression markers and the application for various maize varieties.
서언
옥수수(
Zea mays L.)는 세계적으로 3대 작물 중 하나로써, 식량뿐만 아니라 사료와 산업용 원자재에도 널리 이용되고 있으며, 생육기간 동안 다양한 생물학적 및 비생물학적 스트레스 요인들로 인해 생산 및 수량에 영향을 받는다(
Betrán et al. 2003,
Tollenaar & Lee 2002). 옥수수는 다른 작물들에 비해 키가 큰 편이고 잎이 넓기 때문에 특히 한발에 더욱 치명적이며, 생산성 손실에 가장 중요한 요인 중 하나이다. 또한 한발은 옥수수 유묘기, 개화기 직전, 개화기의 anthesis-silking interval(ASI), 잎의 노화등에 영향을 미칠 수 있다(
Leach et al. 2011).
옥수수의 유전체는 7,000만년 전부터 유전체 일부분의 복제가 이루어 졌다고 알려졌으며, 유전체 전체의 복제는 1,200만년 전부터 500만년 전에 일어났다(
Blanc & Wolfe 2004,
Paterson et al. 2004,
Swigonova et al. 2004). 이로 인해 가깝게 연관되어 있던 작물인 수수로부터 구분되었다(
Paterson et al. 2009). 이로 인해 옥수수 유전체 중 10개의 염색체는 구조적으로 다양해지고, 염색질 조성에서 역동적인 변화를 겪었다. 옥수수 연구에 어려움이 있다면 최근 300만 년간 교배를 통한 long terminal repeat retrotransposons (LTR retrotransposons)의 급격한 증가로 인해 유전체 크기가 타 작물보다 크다는 점이다(
SanMiguel et al. 1998). 대표적인 모델 식물인 애기장대는 유전체의 크기가 125 Mbp데 반해 옥수수는 유전체의 크기는 20배나 더 큰 2,500 Mbp이다(
Arumuganathan & Earle 1991). 이는 벼의 유전체 크기에 6배이기도 하다. 이러한 옥수수의 매우 큰 유전체를 연구하고 내병성 및 내재해성 품종을 개량하기 위한 다양한 마커들을 개발함에 있어 많은 양의 데이터베이스와 선별 마커들이 필요하다(
Myles et al. 2009). 최근에 옥수수 연구의 어려움을 극복하기 위해 nested association mapping (NAM) population을 이용하여 분자 육종 연구가 진행되고 있다(
Yu et al. 2008). NAM population은 B73을 공유하고 있는 교잡종 집단으로, B73의 염기서열 해독은 완료되었으며(
Schnable et al. 2009), B73은 생물학적, 비생물학적 스트레스에 중간 또는 약한 성질을 띄고 있다. 이를 이용하여 많은 스트레스 내성 품종을 육종하기 위해 모본 또는 부본으로 사용되고 있다(
Hao et al. 2010,
Krill et al. 2010).
본 연구는 NCBI의 옥수수 유묘기 한발 관련 microarray 데이터를 분석하여 한발 내성 계통과 감수성 계통의 발현패턴 비교분석을 통해 유전자를 선발하고, 다양한 품종에 적용하여 한발에 대한 내성과 감수성을 선별하는 것을 목표로 한다. 궁극적으로 옥수수 한발 내성 품종 육성을 위하여 유묘기에 한발 내성 품종을 선발할 수 있는 유전자 기반 발현 마커 개발에 목적이 있다.
재료 및 방법
생육 조건 및 한발스트레스 처리
실험재료는 B73, CML103, CML228, CML277, CML322, Ki3, Ki11, M37W, NC358 총 9개의 옥수수 자식 계통을 USDA-ARS에서 분양 받아 사용하였다(
Table 1). 옥수수 계통의 생육을 위해 지름 130 mm, 높이 180 mm, 바닥 80 mm의 포트에 토양은 질석(vermiculite)을 선택하여 종자를 파종하였다. 상토로 사용되는 질석은 공기 및 수분 친화력이 높기 때문에 뿌리샘플을 얻는 작업에서 뿌리에 붙은 토양을 제거하기가 용이하다. 각각의 포트마다 질석을 10 L씩 채웠으며, 복토의 깊이는 4 cm로 하였다. 수분공급은 각 포트마다 매일 1L씩 공급하였으며, 생육환경은 온도 25°C, 습도 40%가 항상 유지되며 16시간 광조건(LED 램프 및 삼파장 램프, 9500 lux), 8시간 암조건이 일정하게 반복되는 식물배양실 안에서 진행하였다.
Table 1List of 9 inbred lines of maize in study.
Table 1
|
Inbred line |
Origin |
|
B73 |
USA, Iowa |
|
CML 103 |
Mexico, Federal District |
|
CML 228 |
Mexico, Federal District |
|
CML 277 |
Mexico, Federal District |
|
CML 322 |
Mexico, Federal District |
|
Ki3 |
Thailand |
|
Ki11 |
Thailand |
|
M37W |
South Africa, KwaZulu-Nata |
|
NC358 |
USA, North Carolina |
옥수수 유묘기(V2 stage, 발아 후 2주)에 우선적으로 대조군 식물체의 잎과 뿌리에서 각각 RNA추출을 진행하였다. 한발 스트레스 처리군은 뿌리가 손상되지 않도록 물을 이용하여 질석을 제거한 뒤 6시간 및 12시간 동안 페이퍼 타올 위에서 한발 처리를 진행하였다. 한발 처리가 끝난 유묘기 옥수수의 잎과 뿌리에서 각각 RNA를 추출하였다. 물을 재공급한 처리군은 6시간, 12시간 동안 한발 처리를 진행한 유묘기 옥수수에 각각 24시간 동안 뿌리가 잠길 정도로 물을 채운 뒤 뿌리 부분이 빛에 노출되지 않도록 페이퍼 타올과 호일을 덮어 수분을 유지하였다. 물 재공급 처리가 끝난 유묘기 옥수수에서도 잎과 뿌리에서 RNA를 추출하였다.
한발 스트레스 처리는 세번 수행되었으며, RNA은 세번의 처리에서 각각 추출되어 cDNA합성 후 qRT-PCR분석에 사용하였다.
RNA 추출 및 cDNA 합성
RNA 추출은 TRIzol Reagent (Life technologies)를 이용하여 추출하였다. 대조군, 한발 처리 및 물 재공급 처리군의 샘플을 액체질소에 동결시킨 후 막자와 막자사발을 이용하여 곱게 분쇄하였다. 분쇄된 뿌리 조직을 2.0 ml tube에 각각 75 mg씩 넣고 TRIzol Reagent를 1 ml을 첨가하여 균질하게 섞어주었으며, 이후 과정은 제조사의 매뉴얼에 따랄 수행되었다. 추출한 RNA는 nano drop (MAESTROGEN)을 이용하여 정량하였다. 정량된 RNA는 PrimeScript™ 1st strand cDNA Synthesis Kit (TaKaRa)를 이용하여 메뉴얼에 따라 cDNA를 합성하였다.
In silico 분석
Co-expression network 분석을 위하여 NCBI GEO에서 약 1,000개의 microarray에 대한 CEL 파일들을 다운로드하였다. 수집된 CEL 파일을 R package affy를 통하여 RMA 방식으로 규격화하여 통합하였으며, 1,000개의 CEL 파일들을 통하여 구축된 microarray 데이터베이스는 R package the weighted correlation network analysis (WGCNA)를 통하여 co-expression network 분석하였다(
Langfelder & Horvath 2008). 내성 타입과 감수성 타입과의 비교 분석을 통해 옥수수에 한발 스트레스 관련 유전자들을 선발하고, 내성 타입과 감수성 타입에서 유전자들의 발현랑을 MeV program (
http://www.tm4.org/mev.html)을 사용하여 분석하였다. 분석된 모듈은 DAVID (
http://david.abcc.ncifcrf.gov/home.jsp)를 이용하여 functional enrichment analysis을 실시하여 발현양상을 heatmap과 유전자의 수를 venn diagram으로 표시하였다. 클러스터링을 통해 내성 타입과 감수성 타입 사이의 consensus한 모듈을 구축하였다.
Primer는 royal blue 모듈에 포함된 유전자들 중 물 재공급 처리군에서 한발 내성타입과 한발 감수성타입 간 발현 차이를 보이는 유전자 상위 30개를 선발하였다(
Table 2). Reference gene으로는 다양한 환경에서도 가장 안정적으로 발현되는 housekeeping gene인 18s rRNA를 선택하였다(
Manoli et al. 2012). 18s rRNA의 forward primer는 5’-CCATCCCTCCGTAGTTAGCTTCT-3’이며, reverse primer는 5’-CCTGTCGGCCAAGGCTATATAC-3’이다. 한발 처리의 확인하기 위해 internal control로
Dehydration Responsive Element Binding proteins(DREB) 유전자를 이용하였다(
Liu et al. 2013). 본 실험에서는 ZmDREB1A의 subgroup인 ZmDREB1.7을 이용하였다. ZmDREB1.7의 forward primer는 5’-CAGCGACTGCTGGCAAAGC-3’이며, reverse primer는 5’-CAGTAGCTCCAGAGCGTCATG-3’이다.
Table 2Information of primer sequences.
Table 2
|
NCBI Accession no. |
Genechip ID |
Forward primer (5’ → 3’) |
Annealing Temp.(°C) |
|
|
Reverse primer (5’ → 3’) |
|
BU050895 |
Zm.12616.1 |
GCTAATTCCGCAGGAGAGAAA |
56 |
|
TGCCAGCCCTTCTTCTTAAC |
|
CO527263 |
Zm.43.1 |
CTTGTTCGCGAGTCCAGTAT |
56 |
|
CAGCAACGTACCTGCAAATC |
|
AI834513 |
Zm.2073.2 |
CGTGGGTTTGGCTTTGTTAC |
56 |
|
CATTCAGAAGCCTCCCACTAAT |
|
BF728598 |
Zm.1769.1 |
AACCAAGATCGAGCGAATCC |
56 |
|
CATGCGGTTTATTGGCATACAA |
|
CK827168 |
Zm.1987.2 |
ACGGCATGGACACAACTAAA |
56 |
|
ACGGTCCAACTCCTTTGATATG |
|
CD978285 |
Zm.2321.4 |
GCGAAAGTAGTGGCAGAGAA |
56 |
|
CATGCAGGTGCAGATGATAGA |
|
AY104757 |
Zm.14392.1 |
CTCAAGGGCCCACTGTATTATG |
60 |
|
CTCCCAACAACATCGGCTATAC |
|
CO524848 |
Zm.18537.1 |
GAGAGGTCGCGTAGATGATTAG |
56 |
|
TAGGTATTAGCTGCCGATGC |
|
AF457983 |
Zm.13389.1 |
GTTCTTTCTTGCTGTTGCTTAGT |
57 |
|
GATTCCACACTCTCTTCCCTTC |
|
CF627469 |
Zm.2139.1 |
CGTGGGTTTGGCTTTGTTAC |
56 |
|
CATTCAGAAGCCTCCCACTAAT |
|
BG841205 |
Zm.2171.1 |
CGTGGGTTTGGCTTTGTTAC |
56 |
|
CATTCAGAAGCCTCCCACTAAT |
|
BM072816 |
Zm.8068.1 |
GAGAGTCAAAGGGAAGGTGAAG |
56 |
|
ACTTGTTGCCTGACGAGAAATA |
|
CF631845 |
Zm.17625.1 |
ACGAAGCTCGTCGAAGAATATG |
56 |
|
TTGCTCACAGGCGAACTAAA |
|
CF037152 |
Zm.15086.1 |
GGGTAACCAGGATGTGTCTTT |
56 |
|
ACCCTGGAGATGGATTCTTATTG |
|
AY108162 |
Zm.4489.1 |
ATGTCACTCATGACGACTTCTAC |
56 |
|
TCCGTCCTTCCTGCATTAAC |
|
AJ606944 |
Zm.16971.1 |
CCTGTCTTTGCTTCCCTCTTAC |
53 |
|
AACGTGCCCAGCTTCTTT |
|
CO520926 |
Zm.19277.1 |
TCTACTATCTGCCCGTCTACAA |
58 |
|
GAAGTAGTGGAAGCACCAGTAG |
|
BM340313 |
Zm.9828.1 |
GATGAACGAGCGAGGATTGA |
56 |
|
CATAGCAAGTATACCGGAGACC |
|
AW927712 |
Zm.2258.1 |
CGTGGGTTTGGCTTTGTTAC |
56 |
|
CATTCAGAAGCCTCCCACTAAT |
|
BM381236 |
Zm.11849.1 |
CGTGTGCTAGAAAGAACCTGTA |
56 |
|
ACAGAACGAAGACGGAACAA |
|
BG836522 |
Zm.1967.1 |
ACGGCATGGACACAACTAAA |
56 |
|
ACGGTCCAACTCCTTTGATATG |
|
CA403577 |
Zm.4270.2 |
CGAGATGCAAGGTACAAGAAGA |
57 |
|
CCTGAACAGGCAGACCATATT |
|
CB351077 |
Zm.14663.1 |
GTCATCTGGTCGAGTGGAAATAA |
59 |
|
GTCACAGATCGAAGAACCCTAAA |
|
CD219246 |
Zm.16973.1 |
CCTGTCTTTGCTTCCCTCTTAC |
53 |
|
AACGTGCCCAGCTTCTTT |
|
BM078934 |
Zm.15119.1 |
GTGCTGTGGAGCAAGGTAAA |
56 |
|
CCTCATCACTGCCAATCTCTTC |
|
AY103786 |
Zm.281.1 |
GCCACCACCGAAGAAGATAAA |
57 |
|
GAAACGTAAGCGGAGGATCTG |
|
CK145306 |
Zm.17181.1 |
GAGCAGAGAGCAGCACATTA |
56 |
|
CATCTGTAGCACCGCAAATTC |
|
AY110813 |
Zm.3423.1 |
GGGAATTATCTCTCAGCTTCCC |
57 |
|
GCTTGGTCTCTGCAGTCTTAT |
|
BM335820 |
Zm.2227.1 |
CGTGGGTTTGGCTTTGTTAC |
56 |
|
CATTCAGAAGCCTCCCACTAAT |
|
AF329371 |
Zm.445.1 |
TTCTGCGATATGGTCCACAAG |
56 |
|
TTCCCAGCGAGTCAGATAGA |
qRT-PCR은 CFX Connect™ Real-Time PCR Detection System (Bio-Rad)을 이용하였다. Real-Time용 pcr tube에 1/4로 희석한 cDNA 1μl, primer 10pmol, RNase free water 8μl, SYBG mix (Enzynomics) 10μl를 넣었다. qRT-PCR의 조건은 95°C에서 10분간 유지시킨 다음, 45 cycle (95°C 10초, 56°C 10초, 72°C 30초)을 진행한 후 melting curve를 65°C에서 95°C까지 진행하여 상대적 발현 값을 측정하였다. Bio-Rad CFX Manager 프로그램을 이용하여 melting curve 분석을 통해 primer dimer생성 여부를 확인하였다. 또한 qRT-PCR 결과 Cq값 및 Cq평균값을 산출했으며, ΔΔCt method[2^ (Ct sample − Ct ctrl.)]를 이용하여 각각의 상대적 발현값을 확인하였다. 세번의 반복을 통해 산출한 데이터를 바탕으로 발현 패턴을 분석하였다.
결과 및 고찰
In silico 분석
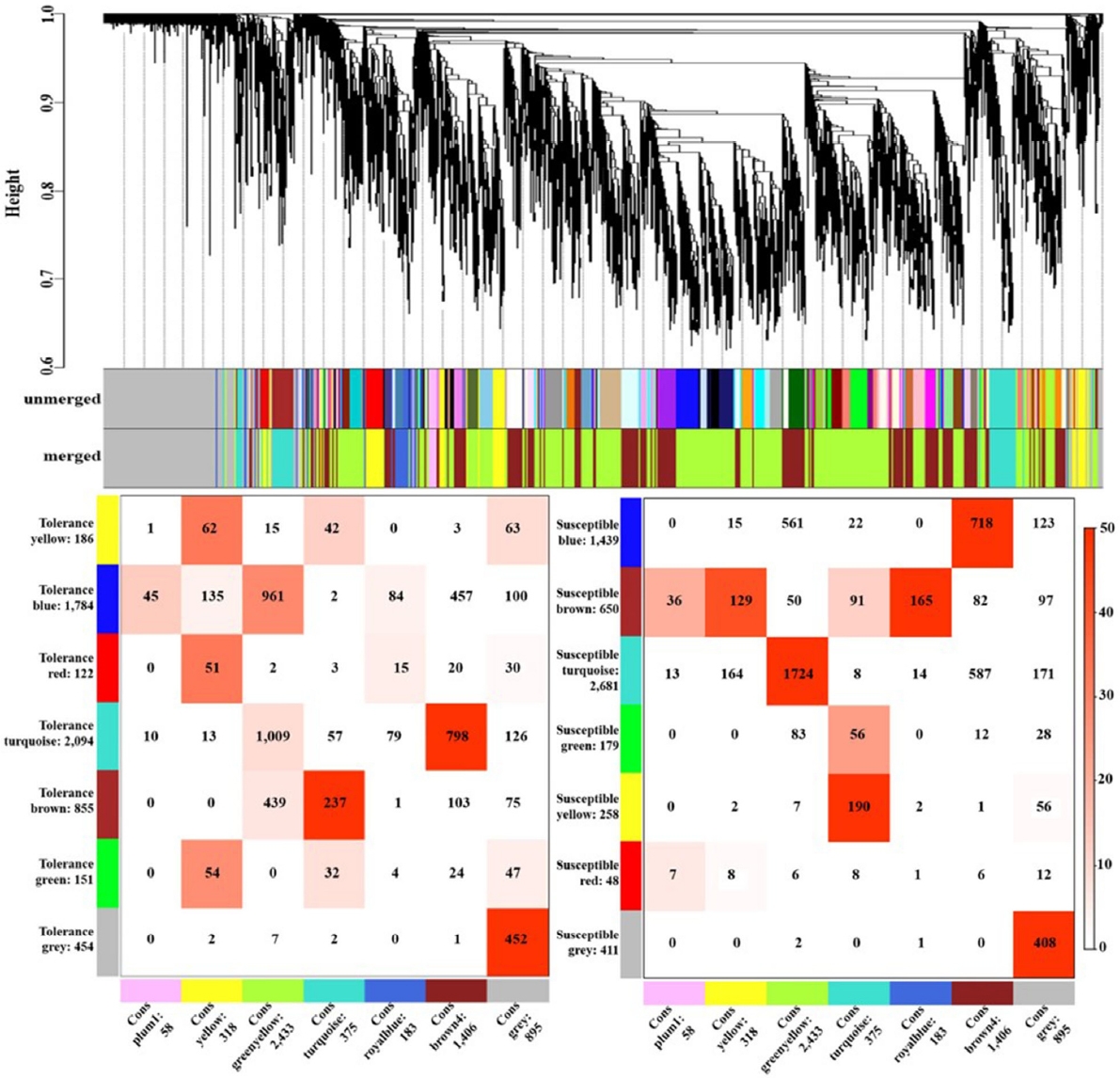
옥수수에서 한발 스트레스에 반응하는 4,771개의 유전자를 바탕으로 6개의 모듈(yellow, green yellow, turquoise, royal blue, brown4, plum1)을 구축하였다. 내성 타입에서는 turquoise, brown5 모듈이, 감수성 타입에서는 green yellow, turquoise, royal blue, brown4 모듈이 drought stress에서 대응관계의 모듈 구조를 보였다(
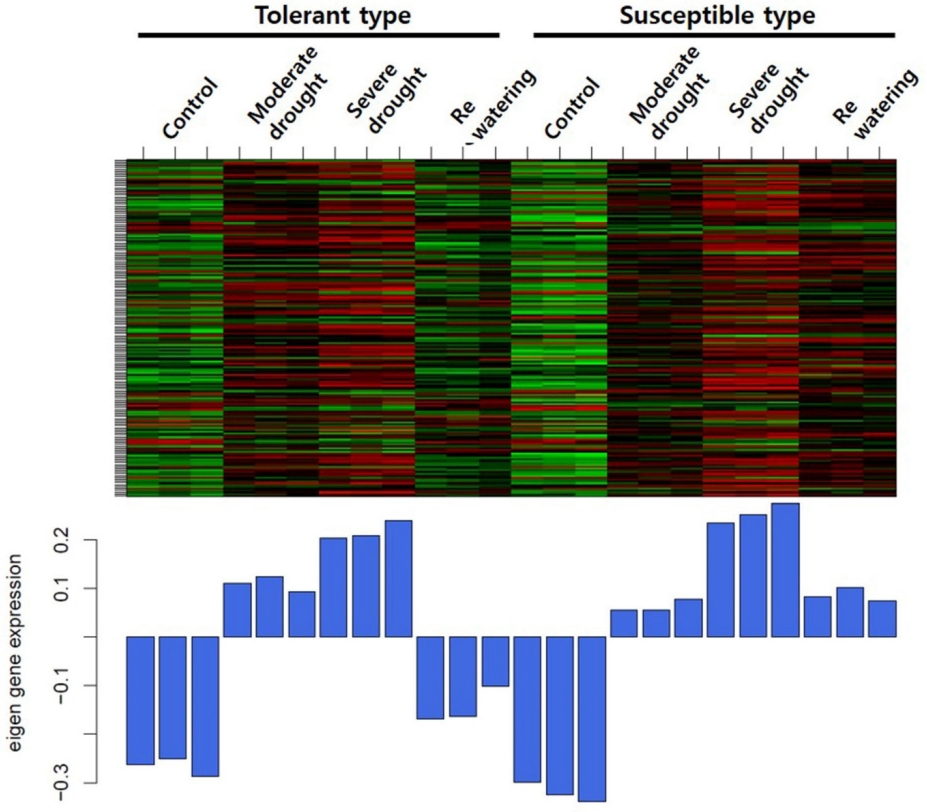
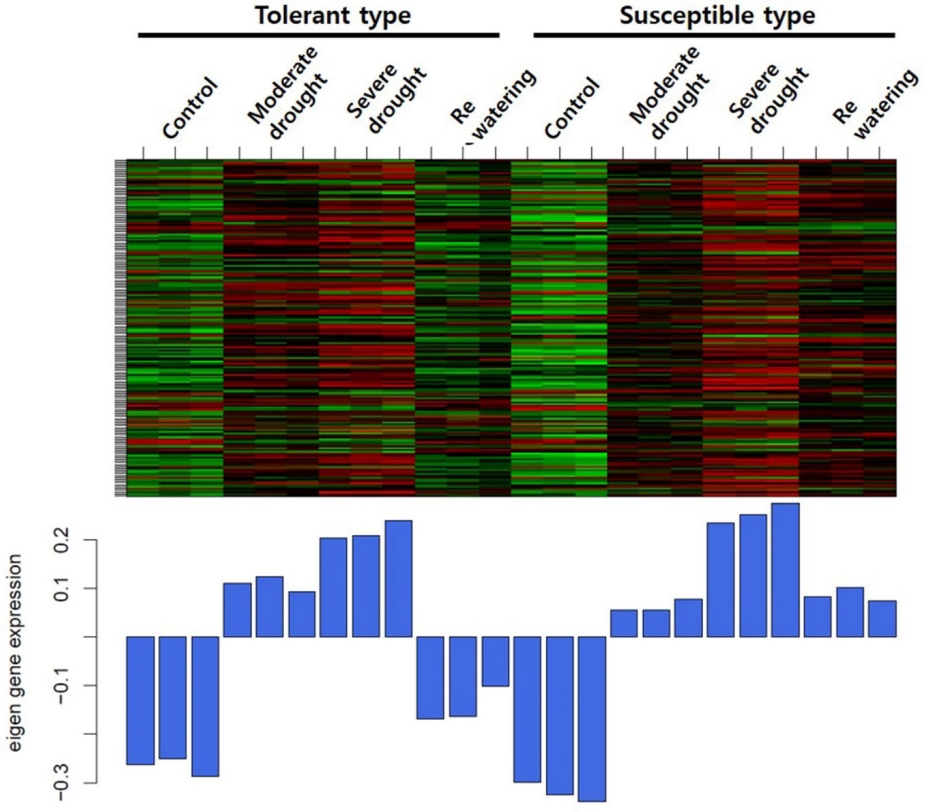
Fig. 1). 클러스터링을 통해 내성 타입과 감수성 타입 간의 consensus한 모듈을 구축하였으며 이를 토대로 클러스터링 트리를 확인하였다. Heatmap을 분석한 결과 6개의 모듈에서 각각 유사한 발현 패턴을 보였고, 모듈별로 yellow 318개, green yellow 2,433개, turquoise 375개, royal blue 183개, brown4 1,405개, 그리고 plum1 56개의 유전자들이 포함되었다. 6개의 모듈 중 royalblue 모듈에서 한발에서는 대조군보다 증가하는 패턴을 보였지만 물 재공급 과정에서 내성 타입에서는 발현이 감소하고 감수성 타입에서는 발현이 증가하는 경향을 보였다. Royal blue 모듈에 포함된 유전자 183개 중에서 110개의 유전자가 내성 및 감수성 타입 모두 물 재공급 처리군에서 발현이 증가하였으며, 45개의 유전자는 감소하는 패턴을 보였다. 또한 4개의 유전자는 내성 타입에서 증가하였고, 24개의 유전자 감수성 타입에서 증가하여 내성 타입과는 반대의 경향을 보였다(
Fig. 2,
Table 3).
Fig. 1Clustering dendrogram of genes and consensus modules found in drought stress-related genes of maize. The correspondence between consensus modules and modules found individually in tolerant and susceptible types based on the expression values of the common genes. Each row corresponds to a drought stress specific module (labeled by color as well as text along with the number of genes in the module), and each column corresponds to one consensus module. Numbers indicate gene counts in the intersection of the corresponding modules. The stronger red color indicates a more significant overlap (left: tolerance type, right: susceptible type).
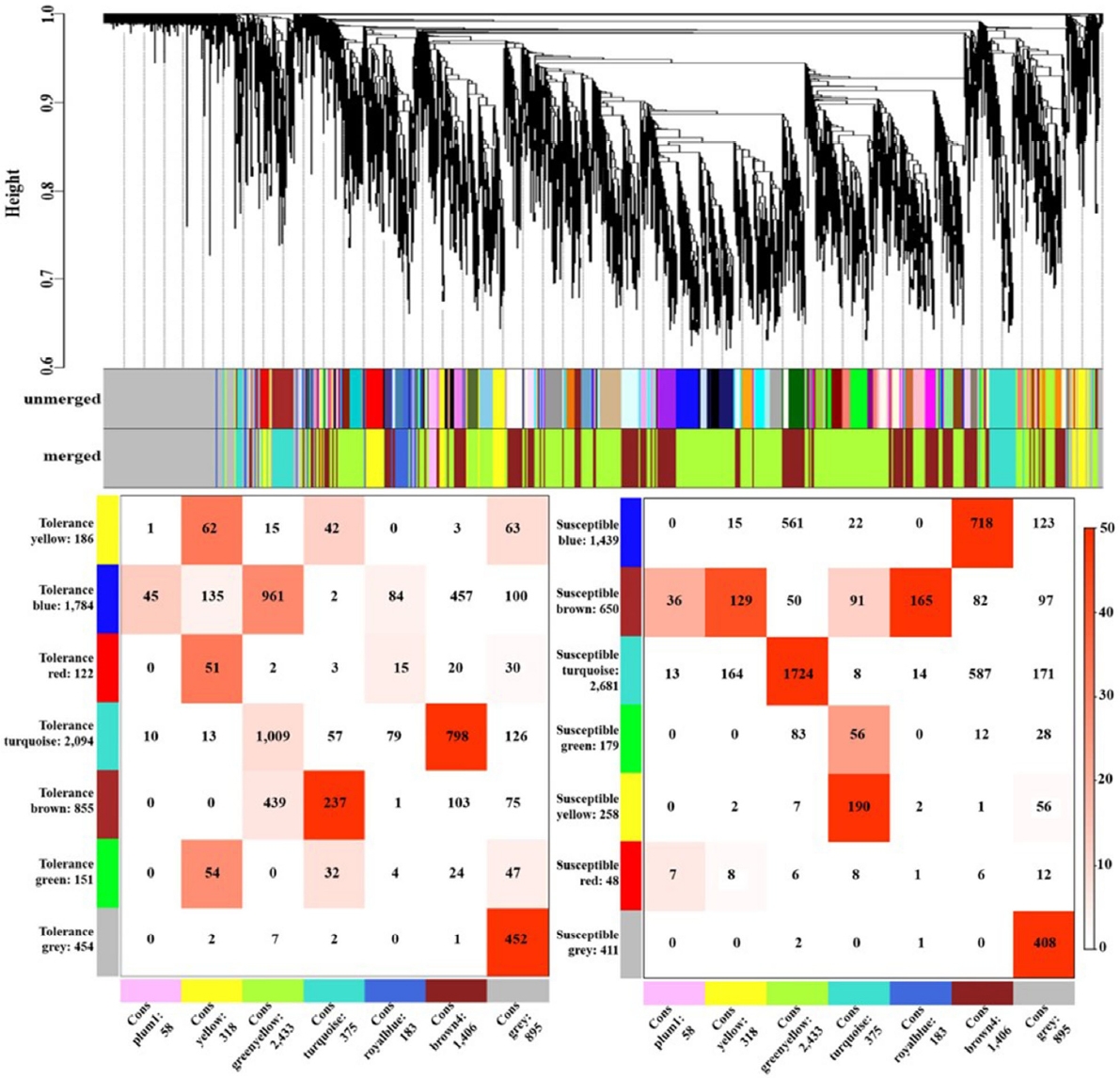
Table 3Number of genes differentially expressed in 6 modules.
Table 3
|
Module |
|
Tolerance type |
Susceptible type |
Overlap |
|
|
MD |
SD |
RW |
MD |
SD |
RW |
MD |
SD |
RW |
|
Yellow |
up |
112 |
88 |
94 |
91 |
90 |
105 |
85 |
87 |
84 |
|
down |
206 |
230 |
224 |
227 |
228 |
213 |
200 |
227 |
203 |
|
Green-yellow |
up |
957 |
974 |
1,269 |
956 |
942 |
1,228 |
880 |
885 |
907 |
|
down |
1,476 |
1,459 |
1,164 |
1,477 |
1,491 |
1,205 |
1,400 |
1,402 |
843 |
|
Turquoise |
up |
191 |
172 |
155 |
198 |
170 |
155 |
130 |
107 |
148 |
|
down |
184 |
203 |
220 |
177 |
205 |
220 |
116 |
140 |
213 |
|
Royalblue |
up |
131 |
131 |
114 |
134 |
134 |
134 |
128 |
128 |
110 |
|
down |
52 |
52 |
69 |
49 |
49 |
49 |
46 |
46 |
45 |
|
Brown4 |
up |
874 |
877 |
625 |
877 |
874 |
698 |
856 |
862 |
356 |
|
down |
532 |
529 |
781 |
529 |
532 |
708 |
511 |
517 |
439 |
|
Plum1 |
up |
27 |
27 |
24 |
28 |
30 |
30 |
26 |
27 |
20 |
|
down |
29 |
29 |
32 |
28 |
26 |
26 |
27 |
26 |
22 |
Fig. 2Heatmap of a royalblue module under drought stress in maize. The x-axis represents microarray samples grouped into tolerant and susceptible types and the y-axis represents genes found in the module. Below the heatmap, the corresponding module eigengene expression values are shown.
qRT-PCR을 이용한 발현양상 분석
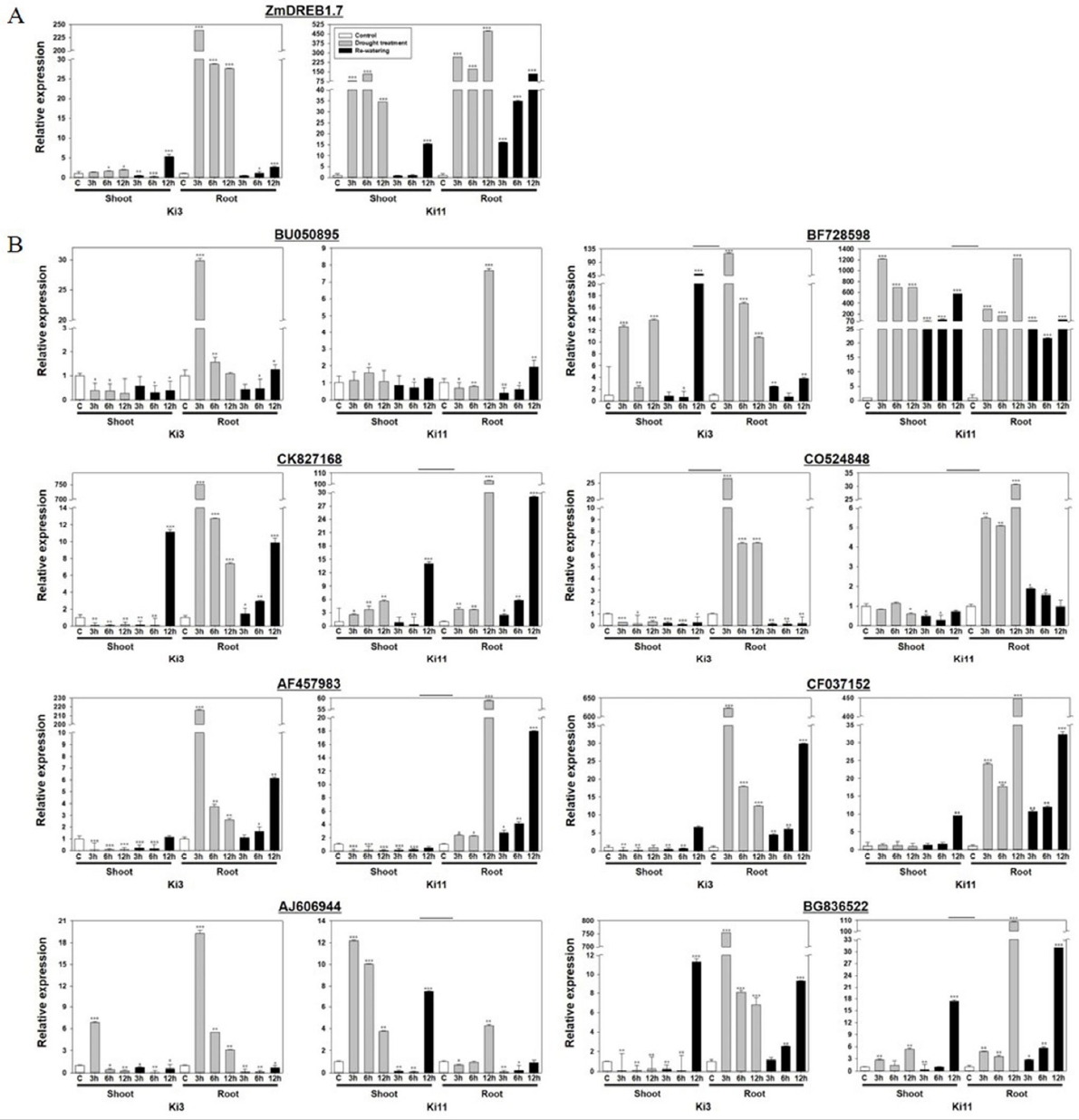
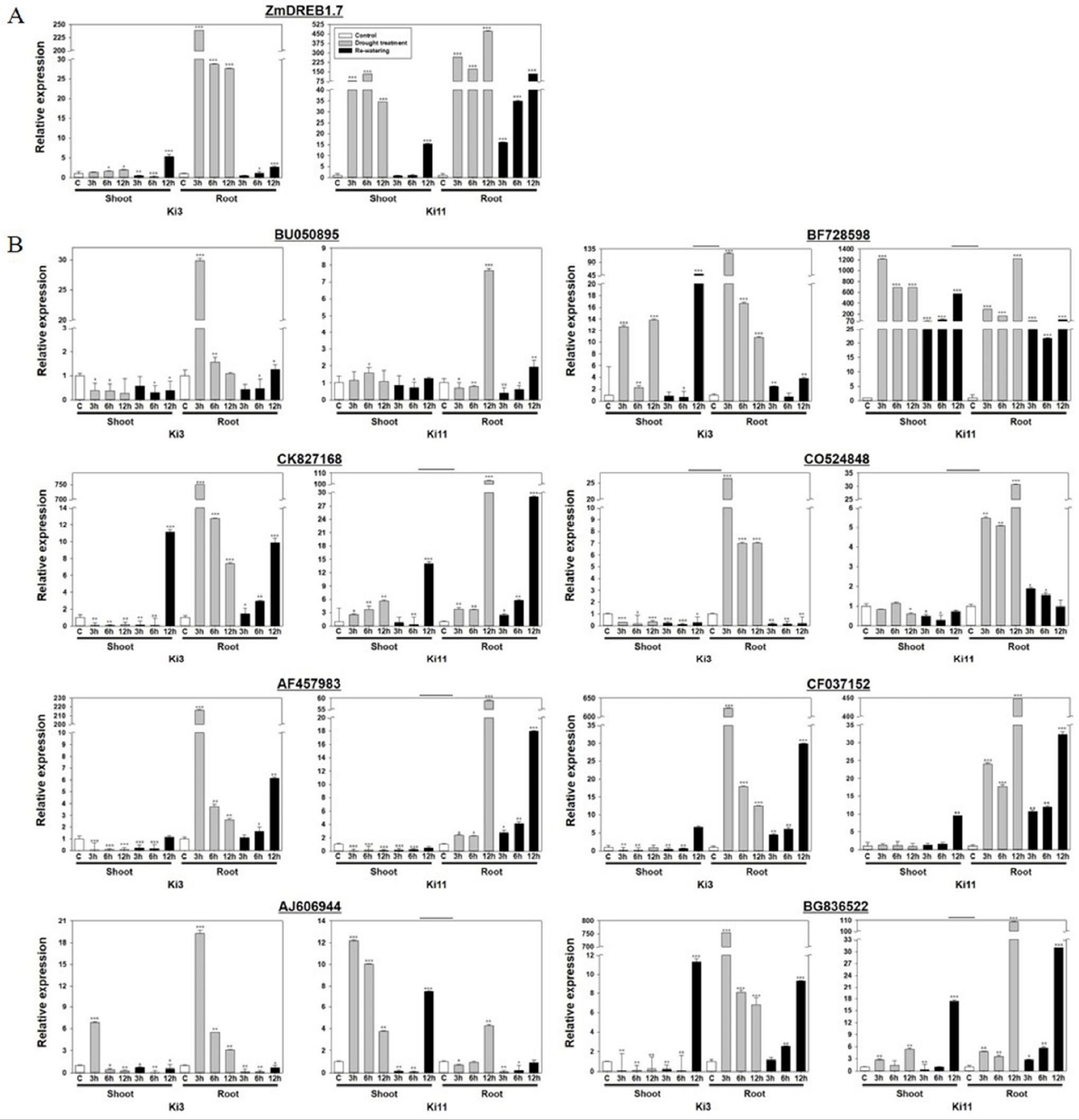
한발 내성타입과 감수성타입 간 발현양상 비교전에 한발 처리가 제대로 수행되었는지 확인하기 위해 ZmDREB1.7의 발현량을 확인하였다. 식물에서 DREB 유전자는 식물에서 수분 스트레스에 관련된 유전자 발현을 조절한다. 옥수수에서 DREB 유전자인 ZmDREB1A와 ZmDREB2A는 수분 스트레스를 받았을 때 발현하게 된다(
Liu et al. 2013). 한발 처리군의 뿌리 부분에서 Ki3와 Ki11은 한발 처리시간이 증가함에 따라 발현량이 증가하였으며, 물 재공급 처리군에서 Ki11은 시간이 증가함에 따라 발현량이 증가한 것을 확인할 수 있었다. 이는 한발 처리된 두 품종 모두 한발 스트레스가 제대로 가해졌음을 확인하였다(
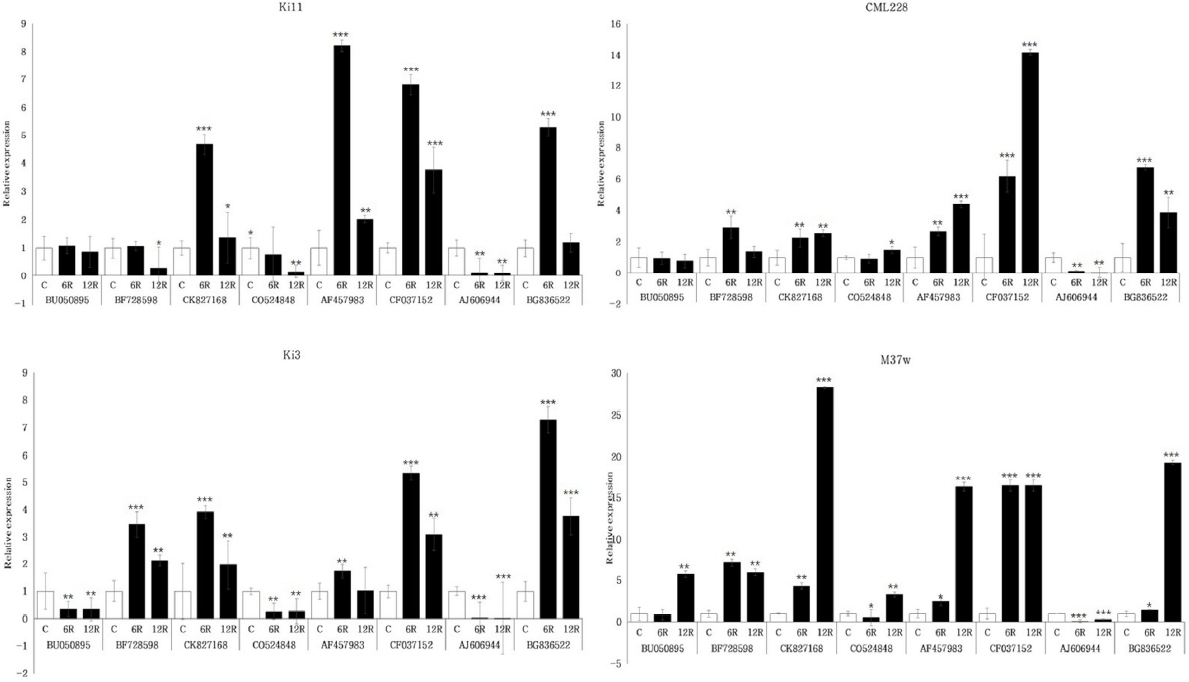
Fig. 3a). 이후 30개 유전자들의 발현 양상을 분석한 결과, BU050895, BF728598, CK827168, CO524848, AF457983, CF037152, AJ606944, BG836522에서 공통적으로 한발 처리군의 뿌리 부분에서 발현 패턴은 처리시간이 증가할수록 Ki3와 Ki11의 발현 패턴이 상반되는 것을 확인할 수 있었다(
Fig. 3b). BF728598과 CO524848의 경우 물 재공급 처리군에서 Ki3와 Ki11에서 다른 패턴을 보이는 것을 확인하였다. 분석 결과를 통해 Ki3와 Ki11의 발현 패턴이 차이를 보이는 유전자 8개(BU050895, BF728598, CK827168, CO524848, AF457983, CF037152, AJ606944, BG836522)를 확인하였고, 그 차이는 주로 뿌리 부분에서 이루어지고 있음을 유추할 수 있었다. 따라서 유전자 30개 중에서 Ki3와 Ki11의 발현 패턴이 차이가 나는 유전자 BU050895, BF728598, CK827168, CO524848, AF457983, CF037152, AJ606944, BG836522를 한발스트레스 관련 발현마커 후보로 선발 하였다.
Fig. 3Expression profiles of Ki3 (susceptible type) and Ki11 (tolerant type) with selected genes under drought stress in maize seedling. A) Internal control (ZmDREB1.7), B) candidate eight genes from selected 30 genes. Data represent the means ±SD of three biological replicates. Asterisks (*, ** and ***) represent significant differences (p<0.05, p<0.01 and p<0.001, respectively) according to the t-test.
후보 유전자의 발현양상 확인
선발된 8개의 유전자를 데이터베이스를 활용하여 기능을 유추하였다(
Table 4). BU050895는 ferredoxin-6으로 엽록체에서 전자전달을 수행하며, 몇몇 연구에서 한발스트레스에서 광합성계의 전자 전달에 영향을 주는 것으로 알려졌다(Lehtimäki et al. 2010,
Mulo 2011). CK827168은 homeobox-leucine zipper HOX24-like로써 세포핵에서 전사인자의 활성과 시퀀스 특이적인 DNA 결합 등을 수행한다. 옥수수에서는 ABA-responsive element, low temperature responsive element 그리고 dehydration responsive element로 알려져 있으며, 한발 스트레스에서 발현이 감소한다(
Zhao et al. 2011). AF457983은 종자 성숙에 역할을 하며, AJ606944는 cytokinin oxidase 3으로 세포 외 공간에서 탈수소 효소의 활성 등을 수행하며, 옥수수의 뿌리에서 다른 조직에 비해 발현이 높고, 사니토키닌의 발현을 조절하고 다양한 비생물학적 스트레스에 반응을 하는 것으로 보고되었다(Brugière et al. 2003). BG836522는 pathogenesis-related 1-like로 생물학적 자극에 반응하는 방어 기작을 수행한다. 유묘기 한발에 강한 계통 4개(CML103, CML228, CML322, Ki11), 유묘기 한발에 약한 계통 2개(Ki3, M37w), 성숙기 한발에 약한 계통 2개(CML277, NC358), reference 품종인 B73에 최종 선발된 유전자들을 적용하여 추가적인 발현 패턴 비교분석을 수행하였다.
Table 4Description of candidate 8 genes from selected 30 genes.
Table 4
|
Gene ID (on B73 RefGen_v3) |
NCBI Accession no. |
Description |
GO Terms |
|
GRMZM2G063126 |
BU050895 |
ferredoxin-6, chloroplastic-like |
chloroplast;electron carrier activity;metal ion binding;2 iron, |
|
- |
BF728598 |
- |
- |
|
GRMZM2G407223 |
CK827168 |
TPA: hypothetical protein ZEAMMB73_259306 |
- |
|
GRMZM2G117164 |
CO524848 |
homeobox-leucine zipper HOX24-like |
chloroplast;electron carrier activity;metal ion binding;2 iron, sequence-specific DNA binding;transcription, DNA-templated;regulation of transcription, DNA-templated |
|
GRMZM2G448627 |
AF457983 |
seed maturation |
embryo development |
|
- |
CF037152 |
- |
- |
|
GRMZM2G167220 |
AJ606944 |
cytokinin oxidase 3 |
extracellular space;cytoplasmic membrane-bounded vesicle; UDP-N-acetylmuramate dehydrogenase activity; cytokinin dehydrogenase activity;flavin adenine dinucleotide binding; |
|
GRMZM2G112538 |
BG836522 |
pathogenesis-related 1-like |
defense response;response to biotic stimulus |
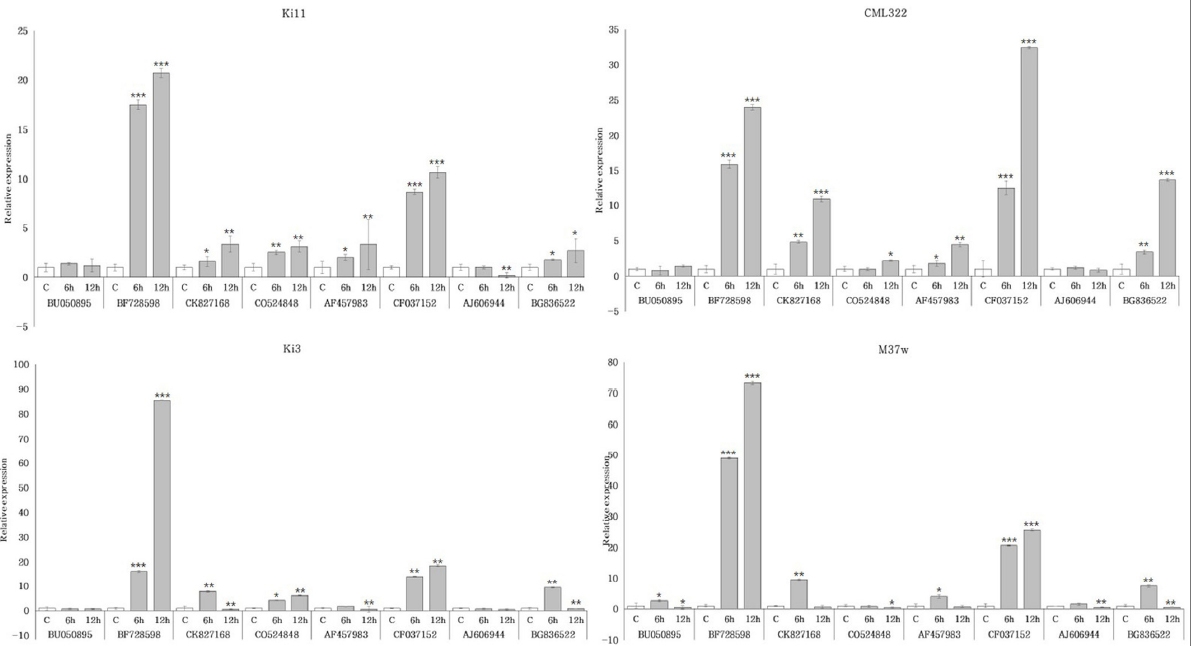
첫 번째로 한발 처리군에서 한발 감수성 타입인 Ki3와 한발 내성 타입인 Ki11간 발현 패턴을 비교한 결과, BF728598, CO524848, CF037152에서 공통적으로 증가하는 패턴의 발현 양상을 보였다(
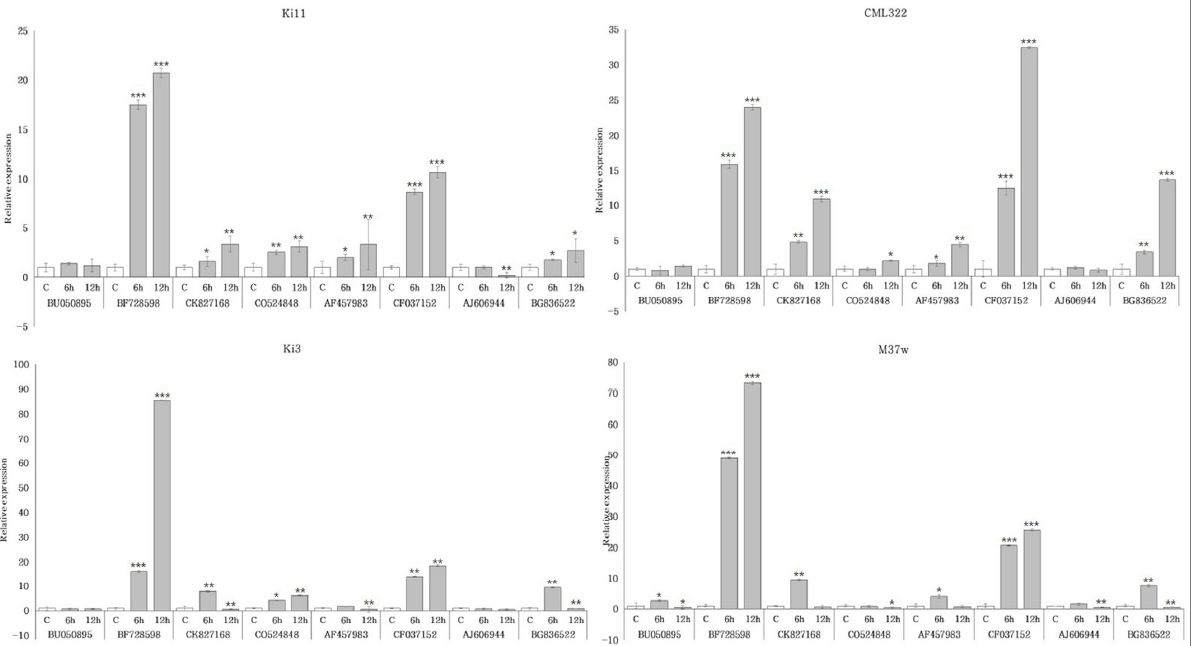
Fig. 4). Ki3는 CK827168, AF457983, BG836522에서 6시간 한발 스트레스 처리군에서 증가하였으나, 이후 12시간 한발 스트레스 처리군에서 감소하는 패턴의 발현 양상이 나타났다. 반면, Ki11은 12시간 한발 스트레스 처리군에서 증가하는 패턴의 발현 양상을 확인함으로써 패턴의 차이가 있는 것을 확인하였다. 유묘기 한발에 강한 품종은 Ki11과 유사하게 BF728598, CO524848, CF037152, CK827168, AF457983, BG836522에서 시간이 지날수록 증가하는 패턴의 발현 양상이 나타났고, 유묘기 한발에 약한 품종에서는 Ki3와 유사한 발현 양상을 보였다. B73은 Ki3와 Ki11의 두 발현 패턴이 모두 나타났으며, 성숙기 한발에 약한 품종은 Ki3와 Ki11과는 다르게 BF728598이 감소하거나 소폭 증가하는 것을 확인하였다.
Fig. 4Expression profiles of four inbred lines with selected candidate eight genes under drought stress in maize root. Ki11 (tolerant type, upper left), CML322 (tolerant type, upper right), Ki3 (susceptible type, lower left), and M37w (susceptible type, lower right) were represented, respectively. Data represent the means ±SD of three biological replicates. Asterisks (*, ** and ***) represent significant differences (p<0.05, p<0.01 and p<0.001, respectively) according to the t-test.
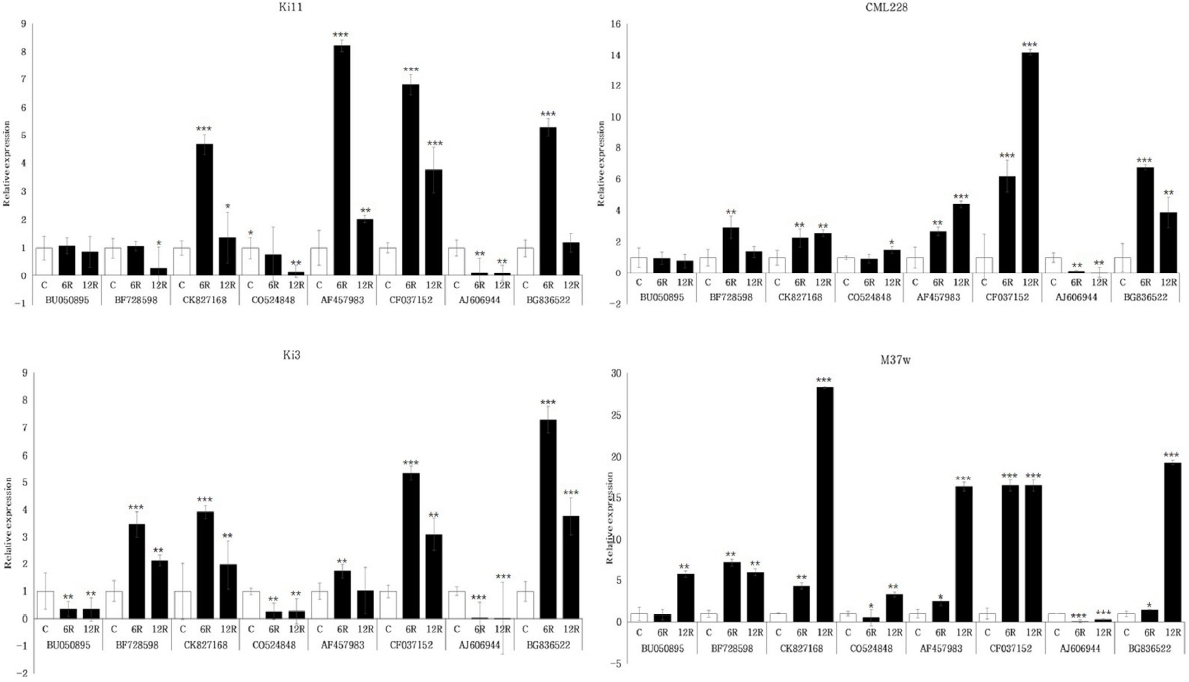
두 번째로 물 재공급 처리군에서 한발 감수성 타입인 Ki3와 한발 내성 타입인 Ki11간 발현 패턴을 비교한 결과, 공통적으로 CK827168, AF457983, CF037152, BG836522는 증가하는 패턴을, BU050895, CO524848, AJ606944는 감소하는 패턴을 확인하였다(
Fig. 5). BF728598에서 Ki3는 증가 이후 유지하는 패턴의 발현 양상을 보였고 Ki11은 시간이 지날수록 감소하는 패턴의 발현 양상을 확인하였다. 유묘기 한발에 강한 품종에서는 Ki11과 유사하지만 BF728598에서 소폭 증가하거나 감소하는 패턴을 보였고, 유묘기 한발에 약한 품종은 Ki3와 유사한 발현 양상을 보였다. B73과 성숙기 한발에 약한 품종에서는 Ki3와 Ki11과는 다른 새로운 패턴의 발현 양상을 확인하였다.
결론적으로, 최종 선발된 8개의 유전자를 이용하여 9개 품종에 적용한 결과 한발 감수성 타입과 내성 타입의 유전자 발현 양상이 다른 것을 확인하였다. 추후에 더 많은 품종들에 적용한다면, 이 8개의 유전자를 이용하여 한발에 감수성인지 내성인지를 유묘기에 판단할 수 있는 한발 관련 발현 마커로써 개발이 가능할 것이다.
Fig. 5Expression profiles of four inbred lines with selected candidate eight genes under rewatering stress in maize root. Ki11 (tolerant type, upper left), CML322 (tolerant type, upper right), Ki3 (susceptible type, lower left), and M37w (susceptible type, lower right) were represented, respectively. Data represent the means ±SD of three biological replicates. Asterisks (*, ** and ***) represent significant differences (p<0.05, p<0.01 and p<0.001, respectively) according to the t-test.
적요
본 연구는 유묘기 옥수수에서 한발 관련 유전자를 이용하여 발현 마커의 탐색 및 개발을 위해 NCBI GEO dataset에서 옥수수 microarray CEL 파일을 이용하여 한발에 관련된 유전자들의 발현을 분석하였다. 유전자들의 co-expression 분석을 통해 6개의 모듈을 구성하였으며, 이 중 한발 내성 타입과 감수성 타입에서 발현 차이를 보이는 royal blue모듈 내의 유전자들 중 상위 30개의 유전자들을 선발하였다. Ki11(한발 내성)과 Ki3(한발 감수성)의 유묘기에 한발 처리 후 qRT-PCR을 이용하여 선발된 유전자들의 발현 패턴을 비교 분석을 통하여 8개의 유전자(BU050895, BF728598, CK827168, CO524848, AF457983, CF037152, AJ606944, BG836522)가 유묘기 옥수수의 뿌리에서 발현 차이를 보이는 것을 확인하였다. 최종 선발된 유전자들을 9개의 품종에 한발 처리 후 뿌리 부분을 타겟으로 qRT-PCR을 진행한 결과, 한발 처리군에서 CK827168과 BG836522가 한발 내성 타입과 감수성 타입 간 발현 양상의 차이를 확인할 수 있었다. 이 결과를 통해 스트레스에 관련된 유전자를 이용하여 발현 마커의 개발 및 적용 가능성을 보여준다.
사사
본 연구는 농림축산식품부, 해양수산부, 농촌진흥청, 산림청 Golden Seed 프로젝트 사업(식량종자사업단, 과제번호: 213009-05-2-SB710)과 농촌진흥청 공동연구사업(과제번호: PJ012649022018)의 지원에 의해 이루어진 것임.
References
- 1. Arumuganathan K, Earle ED. 1991. Nuclear DNA content of some important plant species. Plant Mol Biol Rep 9: 208-218.
- 2. Banziger M, Edmeades GO, Beck DL, Bellon MR. 2000. Breeding for drought and nitrogen stress tolerance in maize:from theory to practice. Mexico CIMMYT.
- 3. Betrán FJ, Beck D, Bänziger M, Edmeades GO. 2003. Genetic analysis of inbred and hybrid grain yield under stress and nonstress environments in tropical maize. Crop Sci 43: 807-817.
- 4. Blanc G, Wolfe KH. 2004. Widespread paleopolyploidy in model plant species inferred from age distributions of duplicate genes. Plant Cell 16: 1667-1678.
- 5. Brugière N, Jiao S, Hantke S, Zinselmeier C, Roessler JA, Niu X, Jones RJ, Habben JE. 2003. Cytokinin Oxidase Gene Expression in Maize Is Localized to the Vasculature, and Is Induced by Cytokinins, Abscisic Acid, and Abiotic Stress. Plant Physiol 132: 1228-1240.
- 6. Buckler ES, Holland JB, Bradbury PJ, Acharya CB, Brown PJ, Browne C, Ersoz E, Flint-Garcia S, Garcia A, Glaubitz JC, et al. 2009. The genetic architecture of maize flowering time. Science 325: 714-718.
- 7. Chardon F, Virlon B, Moreau L, Falque M, Joets J, Decousset L, Murigneux A, Charcosset A. 2004. Genetic architecture of flowering time in maize as inferred from quantitative trait loci meta-analysis and synteny conservation with the rice genome. Genetics 168: 2169-2185.
- 8. De La Cruz M, Romao RL, Escudero A, Maestre FT. 2008. Where do seedlings go? A spatio-temporal analysis of seedling mortality in a semi-arid gypsophyte. Ecography 31: 720-730.
- 9. Fan CY, Hao ZQ, Yan JH, Li GL. 2015. Genome-wide identification and functional analysis of lincRNAs acting as miRNA targets or decoys in maize. BMC Genomics 16:
- 10. Gale MD, Devos KM. 1998. Comparative genetics in the grasses. Proc Natl Acad Sci U S A 95: 1971-1974.
- 11. Gulli M, Salvatori E, Fusaro L, Pellacani C, Manes F, Marmiroli N. 2015. Comparison of drought stress response and gene expression between a GM maize variety and a near-isogenic non-GM variety. PLoS One 10:
- 12. Hao ZF, Li XH, Liu XL, Xie CX, Li MS, Zhang DG, Zhang SH. 2010. Meta-analysis of constitutive and adaptive QTL for drought tolerance in maize. Euphytica 174: 165-177.
- 13. Krill AM, Kirst M, Kochian LV, Buckler ES, Hoekenga OA. 2010. Association and linkage analysis of aluminum tolerance genes in maize. PLoS One 5:
- 14. Langfelder P, Horvath S. 2008. WGCNA:an R package for weighted correlation network analysis. BMC Bioinformatics 9:
- 15. Leach KA, Hejlek LG, Hearne LB, Nguyen HT, Sharp RE, Davis GL. 2011. Primary root elongation rate and abscisic acid levels of maize in response to water stress. Crop Sci 51: 157-172.
- 16. Lehtimäki N, Lintala M, Allahverdiyeva Y, Aro E-M, Mulo P. 2010. Drought stress-induced upregulation of components involved in ferredoxin-dependent cyclic electron transfer. J Plant Physiol 167: 1018-1022.
- 17. Liu SX, Wang XL, Wang HW, Xin HB, Yang XH, Yan JB, Li JS, Tran LSP, Shinozaki K, Yamaguchi-Shinozaki K, Qin F. 2013. Genome-wide analysis of ZmDREB genes and their association with natural variation in drought tolerance at seedling stage ofZea maysL. PLoS Genet 9:
- 18. Manoli A, Sturaro A, Trevisan S, Quaggiotti S, Nonis A. 2012. Evaluation of candidate reference genes for qPCR in maize. J Plant Physiol 169: 807-815.
- 19. Meeks M, Murray SC, Hague S, Hays D. 2013. Measuring maize seedling drought response in search of tolerant germplasm. Agronomy-Basel 3: 135-147.
- 20. Mulo P. 2011. Chloroplast-targeted ferredoxin-NADP+oxidoreductase (FNR):Structure, function and location. Biochim Biophys Acta 1807: 927-934.
- 21. Myles S, Peiffer J, Brown PJ, Ersoz ES, Zhang Z, Costich DE, Buckler ES. 2009. Association mapping:critical considerations shift from genotyping to experimental design. Plant Cell 21: 2194-2202.
- 22. Paterson AH, Bowers JE, Chapman BA. 2004. Ancient polyploidization predating divergence of the cereals, and its consequences for comparative genomics. Proc Natl Acad Sci U S A 101: 9903-9908.
- 23. Paterson AH, Bowers JE, Bruggmann R, Dubchak I, Grimwood J, Gundlach H, Haberer G, Hellsten U, Mitros T, Poliakov A, et al. 2009. The Sorghum bicolor genome and the diversification of grasses. Nature 457: 551-556.
- 24. SanMiguel P, Gaut BS, Tikhonov A, Nakajima Y, Bennetzen JL. 1998. The paleontology of intergene retrotransposons of maize. Nat Genet 20: 43-45.
- 25. Schnable PS, Ware D, Fulton RS, Stein JC, Wei F, Pasternak S, Liang C, Zhang J, Fulton L, Graves TA, et al. 2009. The B73 maize genome:complexity, diversity, and dynamics. Science 326: 1112-1115.
- 26. Swigonova Z, Lai JS, Ma JX, Ramakrishna W, Llaca V, Bennetzen JL, Messing J. 2004. Close split of sorghum and maize genome progenitors. Genome Res 14: 1916-1923.
- 27. Thirunavukkarasu N, Hossain F, Mohan S, Shiriga K, Mittal S, Sharma R, Singh RK, Gupta HS. 2013. Genome-wide expression of transcriptomes and their co-expression pattern in subtropical maize (Zea maysL.) under waterlogging stress. PLoS One 8:
- 28. Thornsberry JM, Goodman MM, Doebley J, Kresovich S, Nielsen D, Buckler ESt. 2001. Dwarf8 polymorphisms associate with variation in flowering time. Nat Genet 28: 286-289.
- 29. Tollenaar M, Lee EA. 2002. Yield potential, yield stability and stress tolerance in maize. Field Crops Res 75: 161-169.
- 30. Unterseer S, Bauer E, Haberer G, Seidel M, Knaak C, Ouzunova M, Meitinger T, Strom TM, Fries R, Pausch H, et al. 2014. A powerful tool for genome analysis in maize:development and evaluation of the high density 600 k SNP genotyping array. BMC Genomics 15:
- 31. Wang L, Liu K, Mao SS, Li ZY, Lu YL, Wang JR, Liu YX, Wei YM, Zheng YL. 2015. Large-scale screening for Aegilops tauschii tolerant genotypes to phosphorus deficiency at seedling stage. Euphytica 204: 571-586.
- 32. Yu J, Holland JB, McMullen MD, Buckler ES. 2008. Genetic design and statistical power of nested association mapping in maize. Genetics 178: 539-551.
- 33. Zhang W, Han ZX, Guo QL, Liu Y, Zheng YX, Wu FL, Jin WB. 2014. Identification of maize long non-coding RNAs responsive to drought stress. PLoS One 9:
- 34. Zhao Y, Zhou Y, Jiang H, Li X, Gan D, Peng X, Zhu S, Cheng B. 2011. Systematic Analysis of Sequences and Expression Patterns of Drought-Responsive Members of the HD-Zip Gene Family in Maize. PLoS One 6: e28488