적요
Salt stress is one of major restrictions for the production of lettuce. In order to identify promising lettuce genotypes having tolerance against salt stress, effect of different sodium chloride (NaCl) concentrations (0, 100, and 200 mM) on electrolyte leakage and growth of thirty-two lettuce landraces from Korea was evaluated. Screening salt tolerant genotypes based on cluster analysis using data of electrolyte leakage discriminated 'IT105183' and 'IT195057' as the most salt tolerant landraces. More importantly, salinity significantly reduced growth of lettuce, but the reduction rates of growth caused by salt stress in salt-tolerant genotypes were much smaller than those in salt-sensitive genotypes. These results indicate that 'IT105183' and 'IT195057' have high potential for being used as parents to improve salt tolerance in other lettuce cultivars.
서 언
국내에서 상추는 엽채류 중 배추 다음으로 많이 생산되고 있으며, 생채로서는 가장 많이 소비되고 있는 채소이다. 2015년 기준으로 상추의 재배면적은 4,022ha이며, 이 중 77%가 시설에 서 재배되고 있다(
Korean Statistics Information Service 2015). 시설재배는 상추의 연중 생산과 출하시기를 조절할 수 있는 장점이 있지만, 시설 내부에서의 강우의 차단과 연중생산과 다수 확을 위한 연작 및 시비 방법 등은 토양에서의 염류집적을 야기하 고 있다(
Shon et al. 1996). 내염성이 약한 식물로 분류되고 있는 상추를 재배하는 동안 염류집적에 의해서 발생하게 되는 토양 내의 높은 염 농도는 광합성의 효율뿐만 아니라 각종 이온의 흡수와 수송을 감소시켜 상추에서 생산성 저하를 초래하고 있다 (
Buwalda & Warmenhoven 1999,
Jin et al. 2004). 염류집적을 억제하기 위해서 녹비작물의 재배, 심토반전, 유공 암거 배수관 설치, 미생물제제 처리와 같은 다양한 제염기술들이 제안되어 왔지만, 현재 농가에서 실용적으로 적용되고 있는 기술은 없는 실정이다(
Jo et al. 2012,
Kim et al. 2013). 따라서 시설재배에서 상추를 재배하는 동안 염류직접에 대한 피해를 억제하기 위한 가장 기본적인 방법은 저항성 품종을 육성하여 재배하는 것이다.
식물은 일반적으로 염 스트레스에 의한 피해를 최소화 시킬 수 있는 내성 기작을 보유하고 있으며, 이러한 능력은 식물의 양적 형질로서 종간 혹은 종내에서 유전 형질에 따라 다양한 차이를 나타내고 있는 것으로 알려져 있다(
Gupta & Huang 2014). 농작물의 재래종 자원은 어떤 특정한 지역에서 오랫동안 재배되어 왔기 때문에 지역 환경 조건에 잘 적응할 뿐만 아니라 그 지역 주민들의 기호도를 충족시킬 수 있는 여지가 크다(
Park et al. 2008). 따라서 재래종 자원에서 내염성이 강한 계통을 선발하여 육종소재로서 활용할 수 있다면, 국내의 재배 농가뿐만 아니라 소비자들의 요구를 충족시킬 수 있는 품종의 육성이 용이할 수 있다. 하지만, 현재까지 국내에서 내염성 상추 품종의 육성을 전제로 하여 국내에서 수집된 재래종 상추에 대한 내염성 평가는 전무한 실정이다. 따라서 본 연구에서는 내염성이 높은 상추 개발을 위한 기초소재를 찾는 것으로 목적으로, 국내 재래종 상추 유전자원을 대상으로 염 스트레스 처리에 따른 생리 및 생장 특성 변화를 검정 하고자 하였다.
재료 및 방법
공시재료
본 연구는 농촌진흥청 유전자원센터에서 보존하고 있는 국내 재래종 상추 32계통을 분양 받아 강원대학교 농업생명과학대학 내의 growth chamber를 이용하여 수행하였다. 분양 받은 각 계통의 종자는 원예용 상토(Seoul Bio, Eumseong, Korea)로 채워진 50구 트레이(Bumnong, Gyeongju, Korea)에 파종하였 으며, 1주일 후 균일하게 자란 실생 개체를 처리 구 당 20개체씩 선발하여 원예용 상토로 채워진 지름 15cm, 높이 11cm 규격의 비닐 포트(Geumsung Chemical, Jeongeup, Korea)에 옮겼다. 정식 2주일 후에 각 식물체에 0, 100, 200mM 농도의 NaCl(Daejung, Siheung, Korea)을 3일 간격으로 300ml씩 공급 하였으며, NaCl 처리 12일 후에 몇 가지 생리 및 생육 변화들에 대해서 검정하였다. 본 실험을 위한 생육 온도, 광도 조건 그리고 일장은 각각 20°C, 1,500lux, 16시간이었다.
NaCl 처리 조건에서의 전해질 누출 함량 및 엽록소 함량 검정
NaCl 처리 12일 후에 각 계통 당 3개 식물체의 제 2엽을 채취하여 탈 이온수로 세척한 후 각 잎당 직경 5mm의 구멍을 잎의 중간부분에서 두 개씩 뚫어 전해질 누출 함량의 측정을 위한 시료를 채취하였으며 전해질 누출 함량은
Julkowska et al.(2016)의 방법을 변형하여 측정하였다. 채취 된 잎 절편은 1ml 증류수에 0.01%의 silwet L-77 solution(Van Meeuwen Chemicals, Weesp, The Netherlands)이 담긴 2ml 튜브에서 담근 후 100rpm으로 2시간 동안 교반하였으며, 전기전도계를 이용하여 1차 전기전도도를 측정하였다. 이후 시료가 담긴 튜브 를 고압증기멸균기에 넣어 121°C에서 20분간 가열하여 조직을 완전히 파괴시켰으며, 100rpm 속도로 2시간을 추가로 교반한 후에 시료의 2차 전기전도도를 측정하였다. 전해질 누출은 1차로 측정한 전기전도도를 2차로 측정한 전기전도도로 나눈 후 100을 곱하여 %로 산출해 내었다. 엽록소 함량은 염 스트레스가 가해진 12일 후에 chlorophyll meter(Minolta, Tokyo, Japan)를 이용하 여 각 개체 당 제 1엽의 잎 중간을 좌우 대칭을 측정한 SPAD 평균값을 활용하여 나타내었다. 전해질 누출과 엽록소 함량은 3반복으로 측정하였다.
염 스트레스 조건에서 생육상의 변화양상은 잎의 면적과 수 그리고 생체중과 건조중의 측정을 통해서 검정하였다. 잎의 면적 은 제1엽을 채취한 후 Delta-T(Devices Ltd., Hertfordshire, UK) 를 이용하여 측정하였으며 잎의 수는 NaCl 처리 12일 후의 전체 잎의 수를 조사하여 나타내었다. 잎의 생체중은 잎의 생체중 측정을 위해서 채취 된 시료를 신선한 상태로 유지하면서 무게를 측정하여 나타냈으며, 건물중은 80도의 열풍건조기(Eyela, Tokyo, Japan)에서 3일 동안 건조시킨 후 생체중과 동일한 방식 으로 측정하여 표기하였다. 생육특성과 관련된 모든 형질들은 전해질 누출과 엽록소 함량과 마찬가지로 3반복으로 측정하였다.
통계분석
통계분석을 위한 모든 결과는 각 계통별로 SPSS 프로그램 (IBM, New York, USA)을 이용하여 평균과 표준오차로 표시하 였다. 또한, 조사 대상 계통들의 염 스트레스에 대한 반응성을 기반으로 세부집단을 구분하기 위해서 전해질누출함량 측정 결과를 이용하여 계층적 군집분석을 실시하였다. 군집분석은 이상치(outlier)의 영향을 적게 받는 중심연결법(Centroid linkage method)을 이용하여 수행하였고, 그룹 사이의 유사성 거리는 유클리언 제곱거리(Squared Eucli-dean distances)로 측정하여 나타내었다.
결과 및 고찰
전해질 누출은 식물체가 내․외부적으로 받는 스트레스에 의한 세포막의 손상에 의해 발생하기 때문에(
Sutinen et al. 1992), 전해질 누출 분석법은 식물에서의 내건성, 내한성, 내염성 등을 평가하는데 있어 유용하게 활용되고 있다(
Burgess & Huang 2014,
Heo et al. 2014,
Julkowska et al. 2016). 따라서 본 실험에서도 각 재래종 유전자원의 내염성은 NaCl이 가해진 조건 에서 전해질 누출 함량을 측정함으로써 평가하였다. NaCl이 가해지지 않은 상추들에서의 평균 전해질 누출 함량은 12.5%였 던 반면, 100mM과 200mM의 NaCl이 가해진 식물체들에서의 평균 전해질 누출 함량은 각각 31.9와 45.3%로 조사되어, NaCl 이 처리 농도의 증가와 함께 전해질 누출 함량이 높아지게 되는 것으로 나타났다(Table
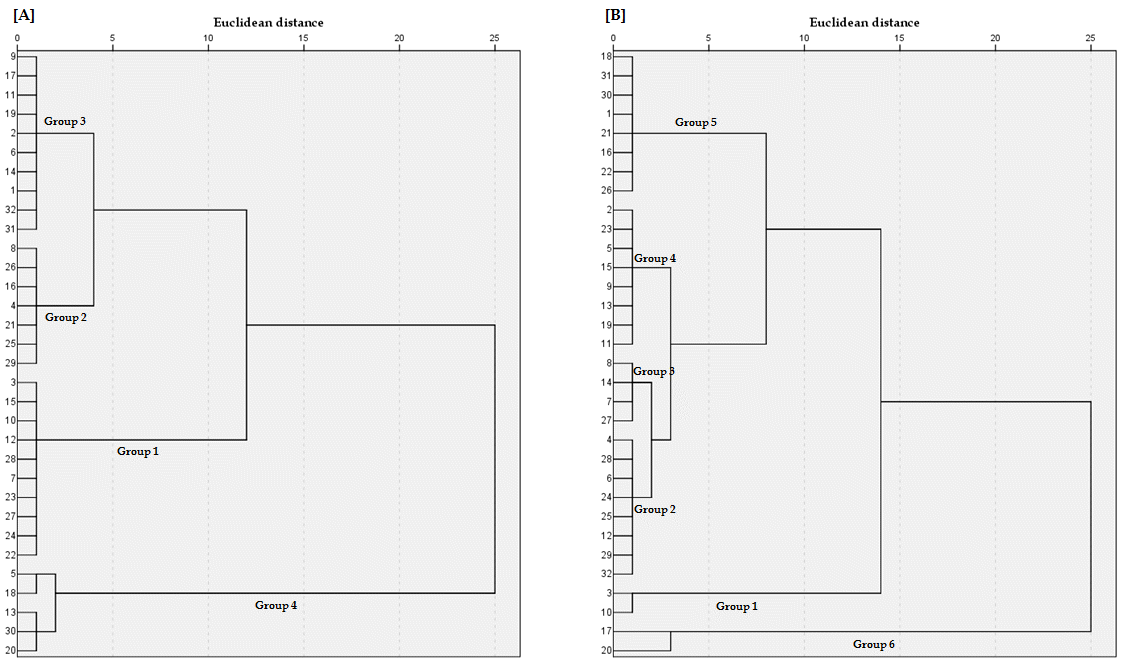
1). 본 실험에 이용된 계통들 간의 내염성 을 구분하고자 군집분석을 실시한 결과 100mM 처리구간에서는 4개의 그룹으로, 200mM처리 구간에서는 6개의 그룹으로 나뉘 게 되는 것으로 나타났다(Fig.
1.). NaCl 처리 조건에 상관없이 IT105183과 IT195057이 제 1그룹에 속하여 본 실험에 이용된 계통 들 중에서 내염성이 가장 강한 것으로, IT218391는 마지막 그룹에 속하여 내염성이 가장 낮은 것으로 조사되었다. 특히 IT235353와 같은 일부의 계통의 경우 낮은 농도의 NaCl 처리 구간에서는 첫 번째 그룹에 속하여 상대적으로 강한 저항성을 가지고 있었지만, 비교적 높은 NaCl처리 구간에서는 다섯 번째 그룹에 속하여 강한 염 스트레스 저항성을 나타내지 못하는 것으로 나타났다. 이러한 양상은 염 스트레스에 대한 치사 농도가 계통에 따라서 다르기 때문에 나타난 결과로 사료되었다. NaCl 처리에 따른 엽록소의 함량의 범위는 100mM의 농도에서는 13.3~40.2, 200mM에서는 16.4~45.9으로써, NaCl 처리가 이루 어지지 않았던 구간(13.2~31.7)에 비해서 엽록소 함량 범위의 차이가 크게 나타나는 것을 확인할 수 있었다. 또한, Table
1에 나타난 것과 같이 대부분의 계통에서 NaCl 처리 농도가 높아짐 에 따라서 엽록소의 함량이 증가하는 양상이 관찰되었다(Table
1). 염 스트레스 조건에서 엽록소 함량의 증가는 피마자, 비트, 상추 등에서 관찰되었으며(
Jamil et al. 2007,
Pinheiro et al. 2008,
Xu & Mou 2015), 이는 엽록체에 염이 축적됨으로써 엽록체의 수가 증가하는 것에 기인하는 것이라고 보고된 바가 있다(
Kirst 1989,
Misra et al. 1997).
Table 1.Effect of different salt concentration on electrolyte leakage, chlorophyll index and fresh weight in lettuce landraces from Korea.
Fig. 1.Dendrogram of the relative tolerance to salinity of 32 lettuce landraces from Korea (A : 100mM salt, B : 200mM salt). On the left side of figure (A), the cluster analysis allowed for grouping the genotypes into four groups: very tolerant (Group 1), tolerant (Group 2), intermediate (Group 3) and sensitive (Group 4). On the right side of figure (B), 32 lettuce landraces from Korea were separated into six groups: very tolerant (Group 1), tolerant (Group 2), upper-intermediate (Group 3), lower-intermediate (Group 4), sensitive (Group 5) and very sensitive (Group 6).
염 처리에 따른 각 재래종 계통 있어 주요 생육 특성의 변화 양상은 Table
2에 나타낸 것과 같다. 식물에서 염 스트레스 반응은 다양한 생리, 화학적 반응의 상호작용에 의해 나타나기 때문에 염 처리 이후 식물에서 나타나는 외형적 특징의 변화만으 로 내염성을 판별하는 것은 쉽지 않은 실정이다. 하지만, NaCl 농도의 증가에 따라서 잎의 면적과 수, 지상부의 생체중과 건물중 의 변화에 있어서 부정적인 연관성이 보고된 바가 있기 때문에 (
Beltagi et al. 2006,
Rui et al. 2009,
Memon et al. 2010), 본 실험에서는 대표 생육 지표로서 지상부의 생체중과 건조중, 잎의 수와 면적의 변화를 측정하여 염 스트레스 조건에서 상추의 반응성을 검정하였다. 본 실험 결과, 염 처리에 따른 생체중의 감소 폭은 매우 큰 반면, 건조중의 감소 폭은 적은 것으로 나타났 다. 각 염 처리 구간 별로 측정 된 생체중의 평균은 100mM와 200mM에서 각각 19.0g과 14.3g으로서 무처리구 대비 15.7%와 36.8%씩 감소하여 전반적으로 고농도의 염 처리 구간에서 감소 의 폭이 큰 것으로 조사되었다(Table
1). 또한, 계통간의 염 스트레스에 따른 생체중의 감소 폭도 고농도의 염 처리구간에서 크게 나타나는 것을 확인할 수 있었다. 예를 들어 내염성이 강한 IT195057와 IT105183에서는 200mM의 염 스트레스 조건에서 생체중이 염을 처리하지 않은 처리구간에 비해서 18.4%와 25.5%씩 감소하여 감소율이 상대적으로 낮았던 반면, 내염성이 약했던 IT218391 계통에서는 49%와 같이 큰 폭의 저하가 일어 나 감소율이 컸다. 염 스트레스는 생체중의 감소뿐만 아니라 상추 잎의 수와 면적의 감소를 야기하는 것으로 나타나 상추의 발육에 있어서도 부정적인 영향을 미치는 것으로 조사되었다 (Table
2). 100mM와 200mM에서의 잎의 수는 염이 처리되지 않은 조건에 비해서 평균적으로 7.8%와 14.2%, 잎의 면적은 17.1와 30.2% 감소하게 되는 것으로 나타났다. 흥미로운 사실은 생체중에서 관찰된 것과 마찬가지로 잎의 수와 면적의 감소는 내염성이 클수록 그 정도가 제한되는 양상을 보였으며, 염 처리 농도가 높아질수록 계통간의 생육저하에 있어서 변이가 커지는 것으로 나타났다는 점이었다(Table
3). 낮은 염스트레스 조건에 서 내염성이 강한 계통들(IT195057와 IT105183)에서의 잎의 수는 5.3%에서 8.6% 범위에서 감소된 반면, 저항성이 약한 계통(IT218391)에서는 16% 가량 감소가 되는 등 그 낙 폭이 상대적으로 크게 나타났다. 이러한 경향은 잎의 면적에서도 거의 동일하여 내염성이 강할수록 잎 면적의 변화가 적어지게 되는 것으로 나타났다. 본 실험의 결과와 유사하게 야생식물에서부터 식용작물까지 광범위한 식물 종에서 염 스트레스가 생육전반의 감소를 야기할 수 있음이 보고된 바가 있다(
Mathur et al. 2006,
Zhao et al. 2007,
Rui et al. 2009). 일부 연구자들에 의해서 토양내에서 염의 농도가 높아질 경우 세포내 삼투압을 위한 수분흡수가 느려지게 됨으로써 기공이 닫히게 되고 세포분열과 신장에 손상이 일어나게 되어 잎 면적의 감소가 나타나게 되고, 이는 결과적으로 원활한 광합성을 저해함으로써 생육 전반에 걸쳐 불량 현상을 야기할 수 있다고 추정된 바 있다(
De Oliveira et al. 2013,
Perez-Lopez et al. 2013). 비록 본 연구에서는 염 처리에 따른 상추계통들에의 수분과 양분 흡수 양상에 대한 검정이 이루어지지 않았지만, 본 실험을 통해서 관찰 된 상추 발육에 있어서 부정적인 변화는 이전 연구들에서 나타난 원인들 에 의해서 발생하게 되는 것으로 사료되었다.
Table 2.Effect of different salt concentration on dry weight, number of leaves and leaf area in lettuce landraces from Korea.
Table 3.Changes of photosynthetic and growth response according to groups segregated by cluster analysis under different salt concentration.
Table 3.
|
100mM salt |
Chlorophyll index (%)z
|
Fresh weight (%) |
Dry weight (%) |
Number of leaves (%) |
Leaf area (%) |
|
|
Group 1 |
13.8ay
|
-13.2a |
-1.5a |
-5.7a |
-13.3a |
|
Group 2 |
24.8a |
-14.2a |
-11.6ab |
-8.6ab |
-18.5ab |
|
Group 3 |
15.6a |
-16.9a |
-8.7ab |
-7.8ab |
-16.1a |
|
Group 4 |
15.7a |
-20.2a |
-14.6b |
-10.5b |
-24.5b |
|
|
200mM salt |
|
|
|
|
|
|
Group 1 |
13.1a |
-21.9a |
2.0a |
-6.9a |
-4.2a |
|
Group 2 |
48.9b |
-36.2bc |
-15.9b |
-14.2bc |
-29.7bc |
|
Group 3 |
36.3ab |
-32.4b |
-15.2b |
-10.9ab |
-18.8b |
|
Group 4 |
37.6ab |
-38.3bc |
-9.0ab |
-14.5bc |
-34.7c |
|
Group 5 |
43.3ab |
-38.2bc |
-21.8b |
-15.3bc |
-35.6c |
|
Group 6 |
15.0a |
-42.0c |
-19.7b |
-16.0c |
-40.6c |
본 실험을 통해서 국내 자생 상추 계통들 간에서 내염성에 대한 유전적 변이가 있음을 확인할 수 있었다. 특히, IT105183과 IT195057 같이 내염성이 강한 계통은 염 스트레스 조건에서도 성장이 큰 폭으로 감소되지 않는 것을 관찰 할 수 있었다. 이상의 연구 결과들을 볼 때, 이러한 계통들은 내염성이 우수한 새로운 품종을 육성하는데 있어서 유용한 유전자원으로 이용 될 수 있을 것으로 판단된다.
적 요
염 스트레스는 상추의 생산량과 품질에 영향을 미치는 중요한 제한요소 중 하나이다. 본 실험에서는 내염성이 높은 상추 개발을 위한 기초소재를 찾는 것으로 목적으로, 32개의 국내 재래종 상추 계통들을 대상으로 몇 가지 농도의 염화나트륨 처리(NaCl 0, 100, 200mM)에 따른 전해질 누출과 생장 특성에 미치는 영향을 평가하였다. 전해질 누출 함량 결과를 사용하여 군집분석 을 실시한 결과 'IT105183'과 'IT195057' 계통이 실험에 이용된 계통들 중 가장 강한 내염성을 가지고 있는 것으로 밝혀졌다. 또한, 염 처리는 상추의 성장을 저해하였지만, 성장의 저해 정도 는 내염성이 강한 계통들이 내염성이 약한 계통들에 비해서 낮은 것으로 나타났다. 따라서, 국내 재래종 상추 계통 중 일부는 내염성이 강한 새로운 상추 품종을 육성하는데 있어 유용하게 이용 될 수 있을 것으로 기대된다.
사 사
본 연구는 강원대학교 대학회계 학술연구조성비로 (과제번호 D1000122-01-01) 수행된 사업입니다.
References
- 1. Beltagi MS, Ismail MA, Mohamed FH. Induced salt tolerance in common bean (Phaseolus vulgaris L) by gamma irradiation. Pak. J. Biol. Sci 2006. 6: 1143-148.
- 2. Burgess P, Huang B. Effects of sequential application of plant growth regulators and osmoregulants on drought tolerance of creeping bentgrass (Agrostis stolonifera). Crop Sci 2014. 54: 837-844.
- 3. Buwalda F, Warmenhoven M. Growth-limiting phosphate nutrition suppresses nitrate accumulation in greenhouse lettuce. J. Exp. Bot 1999. 50: 813-821.
- 4. De Oliveira AB, Alencar NLM, Gomes-Filho E. Akinci S. Comparison between the water and salt stress Effects on Plant Growth and Development. Responses of Organisms to Water Stress. Chapter 4 Rijeka Croatia ISBN 978-953-51-0933-4 2013. InTech Publ. 67-94.
- 5. Gupta B, Huang B. Mechanism of salinity tolerance in plants physiological, biochemical, and molecular chara cterization. Int. J. Genomics 2014. 701596.
- 6. Heo JY, Feng D, Niu X, Mitchell-Olds T, van Tienderen PH, Tomes D, Schranz ME. Identification of quantitative trait loci and a candidate locus for freezing tolerance in controlled and outdoor environments in the overwintering crucifer Boechera stricta. Plant Cell Environ 2014. 37: 2459-2469.
- 7. Jamil M, Lee DB, Jung K, Ashraf M, Lee SH, Rha ES. Effect of salt stress on germination and early seedling growth of four vegetables species. J. Cent. Eur. Agric 2007. 7: 273-281.
- 8. Jin SJ, Cho HJ, Chung JB. Effect of soil salinity on nitrate accumulation of lettuce. Korean. J. Soil Sci. Fert 2004. 37: 91-96.
- 9. Jo SU, Kim DH, Yang JS, Chung KY, Baek KT. Electrokinetic restoration of saline agricultural land. J. Soil & Groundwater Env 2012. 17: 19-26.
- 10. Julkowska MM, Klei K, Fokkens L, Haring MA, Schranz ME, Testerink C. Natural variation in rosette size under salt stress conditions corresponds to developmental differences between Arabidopsis accessions and allelic var iation in the LRR-KISS gene. J. Exp. Bot 2016. 67: 2127-2138.
- 11. Kim SH, Seo DC, Park JH, Lee ST, Lee SW, Kim HC, Cho JS, Heo JS. Effects of green anure crops on growth and yield of carrot for reduction of continuous cropping injury of carrot through crop retation. Korean J Eviron Agric 2013. 32: 279-286.
- 12. Kirst GO. Salinity tolerance of eukaryotic marine algae. Annu. Rev. Plant Physiol. Plant Mol. Biol 1989. 40: 21-53.
- 13. Korean Statistical Information Service. Census of agriculture forestry and fisheries. Seoul Korea 2015.
- 14. Mathur N, Singh J, Bohra S, Bohra A, Vyas A. Biomass production, productivity and physiological changes in moth bean genotypes at different salinity levels. Am. J. Plant Physiol 2006. 1: 210-213.
- 15. Memon SA, Hou X, Wang LJ. Morphological analysis of salt stress response of pak choi EJEAFChe. 2010. 9: pp. 248-254.
- 16. Misra AN, Murmu B, Singh P, Misra M. Growth and proline accumulation in mungbean seedlings as affected by sodium chloride. Biol. Plant 1997. 38: 531-536.
- 17. Park EJ, Kang JH, Kim KH. Characterization of amino acid contents in grain of core collections of Korean native rice. Korean. J. Breed. Sci 2008. 40: 269-277.
- 18. Perez-Lopez U, Miranda-Apodaca J, Munoz-Rueda A, Mena-Petite A. Lettuce production and antioxidant capacity are differentially modified by salt stress and light intensity under ambient and elevated CO2. J. Plant Physiol 2013. 170: 1517-1525.
- 19. Pinheiro HA, Silva JV, Endres L, Dos Santos BG. Leaf gas exchange, chloroplastic pigments and dry matter accumulation in castor bean (Ricinus communis L) seedlings subjected to salt stress conditions. Crop Prod 2008. 27: 385-392.
- 20. Rui L, Wei S, Mu-xiang C, Cheng-jun J, Min W, Bo-ping Y. Leaf anatomical changes of Bruguiera gymnorrhiza seedlings under salt stress. J. Trop. Subtrop. Bot 2009. 17: 169-175.
- 21. Sohn SM, Han DH, Kim YH. Chemical characteristics of soils cultivated by the conventional farming, greenhouse cultivation and organic farming and accumulation of NO3- in Chinese cabbage and lettuce. Korean. J. Org. Agri 1996. 5: 149-165.
- 22. Sutinen ML, Palta JP, Reich PB. Seasonal differences in the freezing stress resistance of needles of Pinus nigra and Pinus resinosa evaluation of the electrolyte leakage method. Tree Physiol 1992. 11: 241-254.
- 23. Xu C, Mou B. Evaluation of lettuce genotypes for salinity tolerance. HortScience 2015. 50: 1441-1446.
- 24. Zhao GQ, Ma BL, Ren CZ. Growth, gas exchange, chlorophyll fluorescence and ion content of naked oat in response to salinity. Crop Sci 2007. 47: 123-131.